Abdominal aortic aneurysm (AAA) and aortoiliac occlusive disease (AIOD) constitute two most common diseases with an indication for elective surgery of the abdominal cavity. Aneurysm is a local widening of the arterial lumen by 50%, as compared to the normal, unchanged part of the artery, right above the aneurysm (1-4). This is a relatively common ailment, found in older individuals mostly. The majority of the cases are asymptomatic. The process of arterial widening follows gradually, over a few years, leading to the rupture of the aneurysm, which often results in patient's demise. The most common location of the aneurysm is the abdominal aorta, right below the renal arteries, but the pathological process may involve other arteries as well.
Mechanical properties of arteries,
i.e. their stretching potential, are conditioned by the presence of two proteins: collagen and elastin. Collagen of the arteries is mostly of the I and III type and is constantly produced by arterial smooth muscles, throughout the whole life. Collagen deficits in the arterial wall may be effectively reversed if the mechanisms of synthesis are efficient and the cause of collagen damage is quickly eliminated. Collagen is a protein that is not susceptible to stretching forces. It is more like a scaffolding of a vessel. The second protein building the vessel is the elastin. This is a stretchable protein which provides vessel elasticity allowing for heart-related changes in the vascular diameter. Elastin is not synthesised in the aortic wall of adults. Its half-time is 70 years, so its amount in the aortic wall decreases with age, which leads to aortic weakening and aneurysm formation, as a consequence of coexistence of many other predisposing factors.
Normal contractility of the abdominal aorta is extremely important in the regulation
of the lumen diameter and blood pressure. The influence of the adrenergic system,
and especially of the
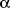 1
1-adrenergic
receptors, is also of significance in this process. The
1-adrenergic
receptors regulate many processes in the brain (all their subtypes), smooth
muscles (
1A
and
1D
-adrenergic subtypes), liver (
1A
and
1B
-adrenergic subtypes), heart, and prostate (
1A-adrenergic
subtype).
The aim of this work was to show the effects of agonists and antagonists of
1-adrenergic receptors on the contractility of aortic smooth muscles within the aneurysmally-changed aorta of humans and the normal aorta obtained from pigs, as well as to show the effects of the abdominal aorta aneurysm on the contractility.
MATERIAL AND METHODS
The experiments were conducted on 18 sections of normal abdominal aortas obtained from pigs weighing from 50 to 70 kg and on 18 sections of aneurysmally-changed abdominal aortas from humans. The human material for the study was collected during elective surgeries of aneurysm correction conducted at the Provincial Specialist Hospital in Wroclaw. Patients from whom the aortic samples were collected, were operated on in an elective or emergent setting, due to an extensive, life-threatening aneurysm of the abdominal aorta. The study was approved by the Ethical Review Board.
The size of aorta sections collected for examination from pigs and from humans,
was the same,
i.e. 0.5x1 cm. The human samples were transported in the
Krebs-Henseleit buffer, at 4°C. Transport time did not exceed 1 hour. Any atherosclerotic
plaque together with the endothelium were removed from the delivered aorta with
the help of a scalpel. The sections were placed horizontally in 4 chambers of
the automatic water bath, with a volume of 20 ml each. Every section was punctured
at its edges with a needle, and a surgical thread safil 4.0 was inserted through
the holes. All the samples were stretched to a tension of 5 mN. That tone was
a baseline used for comparisons of the obtained results (5, 6). The time required
to balance the record was determined experimentally at 20 minutes (7). The Krebs-Henseleit
buffer was used as the incubation environment. It consisted of NaCl - 118 mM,
KCl - 4.7 mM, CaCl
2 - 2.5 mM, MgSO
4
- 1.6 mM, NaHCO
3 - 24.3 mM, KH
2PO
4
- 1.18 mM, and glucose 5.6 mM (5, 7). Incubation of the sections was carried
out at a temperature of 37°C, in the gaseous mixture of oxygen and carbon dioxide
used in the following proportion: 95% of O
2
and 5% of CO
2, in order to obtain the pH value
of 7.3-7.5. Aortic contractions were registered with isotonic transducers Letica
Scientific Instruments) combined with bridge amplifiers (BridgeAmp, ADInstruments,
Australia), a 4-channel data acquisition system (PowerLab/400, ADInstruments)
connected with a Macintosh computer. Spontaneous contractile activity of the
aortic muscle was recorded for 20 minutes (8). Afterwards, agonists and antagonists
of adrenergic receptors were introduced to the incubation chambers with an isolated
section material. The following chemical substances were added: adrenaline -
i.e. agonist of
-
and ß-adrenergic receptors (Sigma-Aldrich), phenylephrine - agonist of
1-adrenergic
receptors (Sigma-Aldrich), prazosine - antagonist of
1-adrenergic
receptors (Sigma-Aldrich), phentolamine - antagonist of
1-
and
2-adrenergic
receptors (Sigma-Aldrich), cyclazocine - antagonist of
1B
receptors (Sigma-Aldrich), L-765314 - antagonist of
1B
receptors (Sigma-Aldrich), BMY 7378 - antagonist of
1D
receptors (Sigma-Aldrich), 5-methylurapidil - antagonist of
1A
receptors (Sigma-Aldrich).
Doses of the preparations were established experimentally by 10-fold dilutions. When defining the experimental dose, we introduced the preparation to the incubation chamber, starting with the highest dilution and not washing the chamber between the administration of subsequent doses (which led to the accumulation of the doses), until a visible effect was obtained on the chart. The experimental dose was defined as the lowest concentration of the preparation, leading to the required effect confirmed in a few subsequent experiments.
The following experimental designs were used in the study:
- an agonist was introduced to the incubation chamber, with dose accumulation
- an antagonist was introduced to the incubation chamber, with dose accumulation
- an antagonist, followed by phenylephrine, was introduced to the incubation chamber, with dose accumulation
Between particular experiments, the incubation chambers were rinsed with Krebs- Henseleit buffer at a temperature of 37°C, three times, and the whole buffer was always removed from the chamber. Only one type of substance was always tested on one section. Before another substance was introduced, the chamber was washed with distilled water and the section was changed for a different one.
When evaluating the obtained results, we analysed the strength of contractions expressed in mN (contractility amplitude). The results of the tests were processed with the use of Microsoft Office Excel 2000 spreadsheets and analysed statistically with Student's t-test and a single-factor analysis of variance (ANOVA) for independent variables.
RESULTS
Sections of the aneurysmally-changed abdominal aorta, obtained from individuals
during elective surgeries of aneurysm correction had a long life-span and could
survive
in vitro for 6-8 hours. The survival time was similar for aortic
sections collected from pigs (no statistically significant differences). Proper
handling of the section material was very important. After sampling, the material
was washed in a cooled Krebs- Henseleit buffer several times, to get rid of
blood and all impurities. The sections had to be transported to the
in vitro
laboratory as soon as possible. Not following the commonly accepted transportation
procedures results in a slow death of the sampled material. After 20 minutes
of control recording (during which the strength of the muscle tone stabilised
and the metabolic activity of the tissue was rising due to temperature and oxygenation
increase), agonists and antagonists of adrenergic receptors were added to the
incubation chamber, in the system of accumulated doses, starting from the lowest
concentration. Sections of porcine abdominal aortas were handled in the same
manner
i.e. stored in a cool buffer for 30 minutes. Both the aneurysmally-changed
abdominal aorta from humans and the normal porcine aorta respond to the administration
of selected agonists and antagonists with muscle tone change. An increase (vasoconstriction)
or a decrease of muscle tone (dilatation) is observed.
Figs. 1-5 show
the obtained results, for a better visualisation of changes and differences
in the contractility of aneurysmally-changed aortas and normal porcine aortas.
 |
Fig. 1. Influence of different
doses of adrenergic antagonists on the contractile activity of smooth
muscle cells of human abdominal aortic smooth muscle. |
 |
Fig. 2. Influence of different
doses of adrenergic antagonists on the contractile activity of smooth
muscle cells of swine abdominal aortic smooth muscle. |
 |
Fig. 3. A comparison of contractile
activity of the aneurysmally-changed abdominal aorta in humans and of
porcine abdominal aorta (control group) in response to the administration
of agonists of -adrenergic
receptors in the system of dose accumulation. |
 |
Fig. 4. Influence of blocking
the subtypes of -adrenergic
receptors with particular agonists and antagonists on the contractile
response of the aneurysmally-changed abdominal aorta in humans to phenylephrine
administration in the system of dose accumulation. |
 |
Fig. 5. Influence of blocking
the subtypes of -adrenergic
receptors with particular agonists and antagonists on the contractile
response of the porcine aorta to phenylephrine administration in the system
of dose accumulation. |
DISCUSSION
The study material comprised of section samples of the abdominal aorta changed by aneurysms and of normal abdominal aortas collected from sows. The human material included sections collected from patients subjected to surgery of aneurismal correction. The age of those patients ranged from 60 to 92 years. Their mean age was 73 years, which proves that aortic aneurysms develop in individuals over 60, this is at least when surgery is needed. Very often, such patients have many comorbidities apart from the aneurysm. In the study group, there were 3 patients with diabetes, 13 with hypertension, 11 with heart diseases, and one with lung and CNS disease. Most of the patients (13 individuals) did not reveal any symptoms of disease connected with the presence of the aneurysm of the abdominal aorta. Three patients were operated on in acute emergency procedures due to aneurismal rupture, and two showed symptoms of pain of the sacrolumbar spine and abdomen. Clot in the aortic lumen was found intraoperatively in all cases; in 4 cases, there was wall thickening and atherosclerotic changes, and in 13 patients there was a thin-walled aneurysm.
The
1-adrenergic
receptor belongs to stimulating receptors, and it is heterogeneous, which means
that it has different subtypes. According to the current classification developed
by the International Union of Pharmacology Subcommittee on Nomenclature for
Adrenoceptors, there are 3 subtypes of
1-adrenergic
receptor:
1A,
1B,
and
1D
(9-15). The
1-adrenergic
receptor is connected with calcium signaling, which means that its stimulation
leads to an increased concentration of intracellular calcium ions (5, 16-18).
The signal is transmitted from the receptor to the cell, by means of such enzymes
as: phospholipase C (PLC), phospholipase D (PLD), and phospholipase A
2
(19, 20). Studies have shown that the effectiveness of
1-adrenergic
receptors in increasing the intracellular concentration of calcium ions is as
follows:
1A>
1B>
1D.
Apart from different subtypes of
1-adrenergic
receptor in humans, there are also nine splice variants of the
1A
-adrenergic receptor,
i.e. 1A-1,
1A-2a,
1A-3a,
1A-4,
1A-2b,
1A-2c,
1A-3c,
1A-5,
1A-6
(10, 21-24).
The experiments showed that the stimulation of the
-adrenergic
receptor causes a contraction of the aortic muscle, both the normal one (swine),
as well as the pathologically changed one (humans). There was a correlation
between tissue reaction and agonist concentration. With increasing concentration,
the intensity of the contraction was increasing as well (
Fig. 3). Aneurysmally-changed
aortas showed a significantly less intense reaction to agonists of a-adrenergic
receptors as compared to porcine aortas (
Fig. 3). The strength of the
contraction is lower. The size of the sections used for examinations was the
same for humans and for swine. This shows that aneurysms inhibit the contractility
and aortic muscle reaction to agonists of
-adrenergic
receptors.
Aneurysms of the abdominal aorta reveal features of a chronic inflammation, destructive reconstruction of the extracellular matrix, decreased number of smooth muscle cells in the tunica media, and a significant reduction of its thickness, as well as neovascularisation (25, 26). As it may be noticed, aneurysms significantly change aortic biomechanics, by influencing the processes of lumen regulation.
The authors distinguished four phases of contraction after -adrenergic agonist administration: 1) phase of a rapid increase, lasting for about 1 minute, with contraction strength of up to 2 g; 2) phase of a slow increase, lasting for 1.5 minute, on average, with contraction strength of up to 3 g; 3) plateau phase lasting for about 4 minutes, with no change in the muscle tone; 4) phase of muscle relaxation, lasting for about 4.5 minutes, with a slow dilatation of the aortic muscle (27, 28). The important factor was the previous removal of the aortic endothelium which significantly reduces muscle contractility and muscle reaction to agonists, which was proved by many authors (7, 27-30). Differences in the contraction strength may also follow from the method of section placement in the incubation chamber (21).
Blockade of
-adrenergic
receptors with phentolamine (nonselective antagonist of a-adrenergic receptor)
results in the lack of contractions in response to adrenaline and phenylephrine
(
Fig. 4, 5). As shown in the study, these are the
-adrenergic
and not the ß-adrenergic receptors that are engaged in the observed vasoconstriction
of the aneurysmally-changed aortic muscle in humans and of the normal porcine
aorta. Administration of phentolamine alone leads to aortic muscle dilatation
(
Figs. 1, 2).
Blockade of that receptor with prazosine (nonselective antagonist of
1-adrenergic
receptor) leads to the absence of tissue reaction to phenylephrine. Administration
of prazosine to the incubation chamber resulted in a gradual dilatation of the
muscle tissue, which confirms the role of
1-adrenergic
receptor in the regulation of aortic vasoconstriction (
Figs. 1, 2, 4, 5).
Maybe the
1-adrenergic
receptor is stimulated
in vitro by agonists - endogenous noradrenaline
secreted in the aortic wall, by postganglionic fibres of the sympathetic system
(31). Similar results were obtained in other animals, when the influence of
phenylephrine on arterial contractility was studied.
In vitro studies
showed that stimulation of
1-adrenergic
receptor by intravenous phenylephrine caused constriction of the arterial vessels
in pigs (32) and dogs (26, 33). The researchers showed a significant increase
in the muscle tone of the arterial vessels, leading to an increased arterial
blood pressure of the examined animals. Blockade of
1A-adrenergic
receptor with 5-methylurapidil resulted in a decrease of the aortic muscle tone
in humans and swine (Figs. 1, 2) and in a decrease of muscle reaction to phenylephrine
Figs. 4, 5).
The results confirmed the presence of
1A-adrenergic
receptors in the aortic muscle and its effects on contractility. and suggested
a key role of this receptor subtype in aortic vasoconstriction caused by the
stimulation of
1A-adrenergic
receptor. The
1A-adrenergic
receptor is not the only receptor influencing the aortic vasoconstriction. Blockade
of
1B-adrenergic
receptors by administration of cyclazocine to the incubation chamber resulted
in a statistically significant decrease in the muscle tone (
Figs. 1, 2).
Blockade of that receptor on the other hand caused a decrease in aortic muscle
reaction to phenylephrine (
Figs. 4, 5). However, the observed changes
are much less pronounced than those registered for the
1A-adrenergic
subtype. Analogous results were obtained with a blockade of
1B-adrenergic
receptor by addition of L765314- selective antagonist of
1B
receptor and blockade of
1D-adrenergic
receptor by administration of BMY 7378 to the incubation chamber. (
Figs.
1, 2, 4, 5).
The literature showed the influence of subtypes of
1-
adrenergic receptor on blood vessels in animals. According to Willems
et
al. (32), phenylephrine influences the contractility of the carotid aorta
in dogs, especially by means of
1A
and
1D
adrenergic receptors; the
1B
subtype is less significant for aortic contractility. The authors showed that
blocking the
1A-adrenergic
receptor in dogs by administering 5-methylurapidil and BMY 7378 at a dose of
100 µg/kg intravenously, reduced the response to phenylephrine by approx. -20±8%,
and a dose of 300 µg/kg completely blocked the response. Abound
et al.
(21) showed that the administration of 5-methylurapidil, benoxanthian and WB
4101 to the incubation chamber decreased the aorta contraction strength in rats
in response to noradrenaline. Blockade of
1B-adrenergic
receptor by addition of L-765314 at a dose of 1000 µg/kg did not have any influence
on tissue reaction to phenylephrine (26, 34, 35). The role of
1B-adrenergic
receptor in aortic vasoconstriction in dogs seems controversial. On the other
hand, this receptor seems to have a crucial role in contractions of the spleen
and splenic vessels, leading to blood expel from the spleen to the peripheral
vessels (16). Administration of BMY 7378 blocked the
1D-adrenergic
receptor in the aorta of the rats, leading to the dilatation (30, 36). However,
its influence on vasa deferentia contractions was not that significant (37).
There are different theories on the mutual functional coexistence of
1A
and
1D
adrenergic receptors in the regulation of vasoconstriction of the renal arteries
in rats (30). Subtypes
1B
and
2B,
on the other hand, did not show any effects on the regulation of contractility
of the arterial vessels in dogs (32) but they took part in regulating the contractions
of the resistance arteries of the skin in rabbits (
1B)
(38) and renal arteries in rats (
2B)
(11).
The presented results as well as literature data confirm the influence of
1-adrenergic receptors on the aortic contractility and show a considerably lower contractile ability of the aneurysmally-changed tissue due to stimulation of
1-adrenergic receptor, as compared to a normal abdominal aorta. This phenomenon seems to be connected with histological changes in the aortic wall, accompanying aneurysm formation (20, 33). The observed loss of elastin and collagen, apoptosis of myocytes of smooth muscles and degradation of the tunica media in the aneurysmally-changed vessel not only inhibit the contractile and elastic function of the abdominal aorta in humans, but also change the reaction of that tissue to stimulation of
1-adrenergic receptors.
Acknowledgments:
This publication is a part of the project "Wrovasc - Integrated Cardiovascular
Centre", co-financed by the European Regional Development Fund, within the Innovative
Economy Operational Program, 2007-2013; "European Funds - for the development
of innovative economy"
Conflict of interests: None declared.
REFERENCES
- Ailawadi G, Eliason JL, Upchurch GR, Arbor A. Current concepts in the
pathogenesis of abdominal aortic aneurysm. J Vasc Surg 2003; 38: 584-588.
- Noszczyk W. Surgery of peripheral arteries and veins. Warszawa, PZWL,
1998.
- Thompson RW, Geraghty PJ, Lee JK. Abdominal aortic aneurysms: basic mechanisms
and clinical implications. Curr Probl Surg 2002; 39: 110-230.
- Wasseff M, Baxter BT, Chisholm RL, et al. Pathogenesis of abdominal
aortic aneurysm: a multidisciplinary research program supported by the National
Heart, Lung and Blood Institute. J Vasc Surg 2001; 34: 730-738.
- Eckert RE, Karsten AJ, Utz J, Ziegler M. Regulation of renal artery smooth
muscle tone by 1-adrenoceptors:
role of voltage-gated calcium channels and intracellular calcium stores.
Urol Res 2000; 28: 122-127.
- Low AM, Darby PJ, Kwan CY, Daniel EE. Efects of thapsigargin and ryanodine
on vascular contractility: cross-talk between sarcoplasmic reticulum and
plasmalemma. Eur J Pharmacol 1993; 230: 6153-6214.
- Caryl EH, Joyce E, Shaun LS. Mechanisms underlying spontaneous rhythmical
contractions in irideal arterioles of the rat. J Physiol 1999; 521: 507-516.
- Gnus J, Czerski A, Zawadzki W, et al. in vitro contractility
of normal and aneurysmal abdominal aorta muscle coat sections in human and
animals material. Folia Biologica (Cracow) 2012; 60: 71-77.
- Brodde OE, Michel MC. Adrenergic and muscarinic receptors in the human
heart. Pharmacol Rev 1999; 51: 651-690.
- Bylund DB, Eilenberg DC, Heible JP, et al. International Union
of Pharmacology nomenclature of adrenoceptors. Pharmacol Rev 1994; 46: 121-136.
- Docherty JR. Subtypes of functional 1-
and 2-adrenoceptors.
Eur J Pharmacol 1998; 361: 1-15.
- Hieble JP, Bondinell WE, Ruffolo RJ. -
and -adrenoceptors:
from the gene to the clinic. Molecular biology and adrenoceptor subclassifcation.
J Med Chem 1995; 38: 3415-3444.
- Hieble JP, Ruffolo JR. Subclassification and nomenclature of 1-
and 2-adrenoceptors.
Prog Drug Res 1996; 47: 81-130.
- Langer SZ. History and nomenclature of 1-adrenoceptors.
Eur Urol 1999; 36: 2-6.
- Liang GH, Park S, Kim JA, Choi S, Suh SH. Stimulation of large-conductance
Ca2+-activated K+
channels by the Na+/Ca2+
exchanger inhibitor dichlorobenzamil in cultured human umbilical vein endothelial
cells and mouse aortic smooth muscle cells. J Physiol Pharmacol 2009; 60(1):
143-150.
- Han C, Abel PW, Minneman KP. al-adrenoceptor subtypes linked to different
mechanisms for increasing intracellular Ca2+ in smooth muscle. Nature 1987;
329: 333-335.
- Minneman K. Alpha 1- adrenergic receptor subtypes. Inositol phosphates
and sources of cell calcium. Pharmacol Rev 1988; 40: 87-93.
- Suzuki E, Tsujimoto G, Tamura K, Hashimoto K. Two pharmacologically distinct
alpha 1-adrenoreceptor subtypes in the contraction of rabbit aorta: each
subtype couples with a different Ca2+ signaling
mechanism and plays a different physiological role. Mol Pharmacol 1990;
38: 725-730.
- O'Donnell SR, Wanstall JC. Beta-1 and beta-2 adrenoreceptor-mediated responses
in preparations of pulmonary artery and aorta from young and aged rats.
J Pharmacol Exp Ther 1984; 228: 733-739.
- Walsh MP. Regulation of vascular smooth muscle tone. Can J Physiol Pharmacol
1994; 72: 919-935.
- Aboud R, Shafii M, Docherty JR. Investigation of the subtypes of 1-adrenoceptor
mediating contractions of rat aorta, vas deferens and spleen. Br J Pharmacol
1993; 109: 80- 87.
- Lomasney JW, Cotecchia S, Lorenz W, et al. Molecular cloning and
expression of the cDNA for the 1A-adrenergic
receptor. J Biol Chem 1991; 266: 6365-6369.
- Pecanha FM, Wiggers GA, Briones AM, et al. The role of cyclooxygenase
(COX)-2 derived prostanoids on vasoconstrictor responses to phenylephrine
is increased by exposure to low mercury concentration. J Physiol Pharmacol
2010; 61: 29-36.
- Stephenson JA, Summers RJ. Autoradiographic analysis of receptors on vascular
endothelium. Eur J Pharmacol 1987; 134: 35-41.
- Lopez-Candalez A, Holmes DR, Liao S, Scott MJ, Wickline SA, Thompson RW.
Decreased vascular smooth muscle cell density in medial degeneration of
human abdominal aortic aneurysms. Am J Pathol 1997; 150: 993-1007.
- Willems EW, Valdivia LF, Ramirez-San JE, Saxena PR, Villalon CM.
Pharmacological identifcation of the major subtypes of adrenoceptors involved
in the panine external carotid vasoconstrictor efects of adrenaline and
noradrenaline. Life Sci 2001; 69: 143-153.
- Angus JA, Cocks TM, Sato K. The alpha adrenoreceptors on endothelial cells.
Fed Proc 1986; 45: 2355-2359.
- Furchgott RF. Role of endothelium in responses of vascular smooth muscle.
Ann Rev Pharmacol Toxiocol 1984; 24: 175-179.
- Nilsson H. Interactions between membrane potential and intracellular calcium
concentration in vascular smooth muscle. Acta Physiol Scand 1986; 1664:
559-566.
- Villalobos-Molina R, Lopez-Guerrero JJ, Ibarra M. 1D-
and 1A-adrenoceptors
mediate contraction in rat renal artery. Eur J Pharmacol 1997; 322: 225-227.
- Hoffman BB, Lefkowitz RJ. Catecholamines, sympathomimetic drugs, and adrenergic
receptor antagonists. In: Goodman & Gilman's The Pharmacological Basis of
Therapeutics, JG Hardman, LE Limbird, PB Molinoff (eds), New York, McGraw-Hill
Professional 1996, pp. 199-248.
- Willems EW, Valdivia LF, Saxena PR, Villalon CM. The role of several
1-
and 2-adrenoceptor
subtypes mediating vasoconstriction in the canine external carotid circulation.
Br J Pharmacol 2001; 132: 1292-1298.
- Dashwood M, Jacobs M. Autoradiographic study of the alpha-adrenoceptors
of rat aorta and tail artery. Eur J Pharmacol 1985; 115: 129-135.
- Cotecchia S, Schwinn DA, Randall RR, Lefkowitz RJ, Caron MG, Kobilka BK.
Molecular cloning and expression of the cDNA for the hamster 1-adrenergic
receptor. Proc Natl Acad Sci USA 1988; 85: 7159-7163.
- Schwinn DA, Lomasney JW, Lorenz W, et al. Molecular cloning and
expression of the cDNA for a novel 1-adrenergic
receptor subtype. J Biol Chem 1990; 265: 8183-8189.
- Perez DM, Piascik MT, Graham RM. Solution phase library screening for
the identification of rare clones: Isolation of an 1D-adrenergic
receptor cDNA. Mol Pharmacol 1991; 40: 876-883.
- Vargas HM, Gorman AJ. Vascular 1A
adrenergic receptor subtypes in the regulation of arterial pressure. Life
Sci 1995; 57: 2291-2308.
- Smith K, Macmillan JB, McGrath JC. Investigations of 1-adrenoceptor
subtype mediating vasoconstriction in rabbit cutaneous resistance arteries.
Br J Pharmacol 1997; 122: 825-832.