Surface epithelial cells of gastric mucosa are constantly subjected to injurious factors, such as alcohol, acid, base, hyperosmolar solution, stress, and the major human pathogen
Helicobacter pylori (
Hp) (1-3). It is well known that
Hp is a gram-negative, spiral shaped bacterium which colonizes the human stomach of about 50% of the worlds population.
Hp colonizes mostly epithelium of the gastric lining and may also penetrate mucus, reaching pits of gastric glands (1). Although all
Hp-infected subjects develop gastritis, approximately 80% of these individuals remain asymptomatic.
Hp could cause the gastric duodenal ulcers, gastric cancer and mucosa-associated lymphoid tissues (MALT) lymphomas affecting about 15%, 1% and 0.1% of the population, respectively (4, 5). Accordingly,
Hp was classified in 1994 by the World Health Organization (WHO) as a class I carcinogen.
Hp has created a series of adaptive mechanisms enabling survival and proliferation in acidic environment of stomach (1, 3). The most important mechanisms include: the ability to adhere to host epithelial cells, high enzymatic urease activity produced by
Hp, which provides covering capacity for survival of bacteria through neutralising acidic content in the gastric lumen, as well as the presence of proton pump that removes an excess of hydrogen ion (1, 3). Majority of
Hp infected patients develop inflammation of gastric mucosa by the increase of proinflammatory genes as well as direct modulation of cellular function, resulting in promotion of cell growth and the resistance to apoptosis (1, 4, 7). Inflammation triggered by
Hp induces many mediators and cellular effectors that appear to be involved in carcinogenesis, such as a variety of cytokines, chemokines and growth factors as well as a prominent fall in the expression of tumor suppressor genes in those cells (4). The contribution of
Hp-induced inflammation to the development of gastric cancer should be further elucidated (4).
Many cancers of digestive organs, some of which are caused by infectious agents, are known to arise on a backgroud of chronic inflammation (8). This also includes
Hp-induced gastric cancer (8), in which epithelial cells of gastric mucosa are directly exposed to
Hp and its cytotoxic products. Virulence factors encoded by the cytotoxin associated gene pathogenicity island (cagPAI) include CagA protein injected
via the type IV secretion system (T4SS) into host target cells, thus contributing to the development of gastric cancer (8-11).
Recent studies suggest that also fibroblasts may alter the mRNA expression of structural and cell-cycle associated genes in the presence of
Hp (12). It was reported that the stomach infection of
Hp is associated with increased myofibroblasts abundance along with its increased proliferation and migration (13, 14).
Normal stroma contains few fibroblasts, especially myofibroblasts, but their
number rapidly increase in reactive stroma surrounding inflammatory region and
neoplastic tissue, particullary in the case of progression of stomach cancer.
The origin of myofibroblasts remains controversial although fibroblasts are
considered to be the main precursors of myofibroblasts. The transdifferentiation
of fibroblasts into myofibroblasts is driven by cancer cell-derived cytokines
i. e. TGF-ß (15).
Myofibroblasts differ from fibroblasts with expression of alpha-smooth muscle
actin (
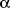
-SMA), a
42kDa molecule, found as a predominant actin protein in aortic smooth muscle
cells (16). The
-SMA
plays a central role in wound contraction, being influenced by various growth
factors and inflammatory cytokines (17). However, in oral wounds that heal without
scar formation, another molecule FGFR1OP2 in fibroblasts is abundant (15). Indeed
-SMA positive myofibroblasts
correlate with enhanced production and deposition of extracellular-matrix related
proteins like collagen type I involved in the process of fibrosis (15, 18).
Myofibroblasts contribute to various steps of the cancer progression including
the increased cell invasion and the suppression of apoptosis (19, 20) as well
as the stimulation of stromal cells with increased production of growth factors
such as HGF that in turn, can affect epithelial cells and enhance various growth
factors (20). The changes in the stroma may drive invasion of cancer cells and
metastasis (20, 21).
In the light of recent studies, heat shock proteins (HSP) are recognized as important family of chaperon proteins involved in cell integrity and defense as well as agents participating in stress response associated with gastric infection of
Hp (22). Among HSP family, in particular heat shock protein-70 (HSP70) seems to be responsible for maintenance of cell integrity under normal cell growth and at the pathophysiological condition such as colonization of gastric mucosa by
Hp (22).
Recently HSP32 (heme oxygenase-1) has been shown to play a significant role in the mechanism of mucosal integrity of upper GI tract (23, 24). However, the role of heme oxygenase-1 (HO-1) in the process of cancer development may depend on the type, location and the degree of advancement of cancer. The upregulation of HO-1 gene exhibits a cytoprotective activity protecting healthy cells against their transformation into the cancer cells. On the other hand, HO-1 through its antiapoptotic and proangiogenic actions has been shown to facilitate the increase in cancer and its invasion. HO-1 may also strengthen cancer cell resistance to radiochemiotherapy, chemo- and fotodynamic therapies (25, 26).
The
Hp-derived reactive oxygen species (ROS) were shown to stabilise
hypoxia inducible factor-1
(HIF-1
), leading
to its increased expression (8). HIF-1
activity appears to be a very early event in carcinogenesis. Previous studies
documented that the mRNA of this protein is increased before the histological
evidence of angiogenesis or cancer cell invasion was confirmed (29). It has
been postulated, that activated HIF-1
is sufficient to functionally confer the cancer-associated fibroblast phenotype
(30). HIF-1
expression
is required for the induction of autophagy in cancer cells. Autophagy in cancer-associated
fibroblasts promotes tumor growth
via the paracrine production of recycled
nutrients, which can directly feed cancer cells (30). HIF-1
activation in different cell types can either promote or repress tumorigenesis
(30, 31). Moreover, fibroblasts harboring activated HIF-1
promote the mitochondrial activity of adjacent cancer cells in a paracrine fashion.
HIF-1
may then
act as a tumor promoter in stromal cells but as a tumor suppressor in cancer
cells (30, 31).
This study, was designed to determine the effect of
Hp on the rat gastric
fibroblasts
in vitro and to examine the influence of
Hp on markers
of transition of fibroblasts into myofibroblasts and extracellular matrix proteins,
-SMA and collagen
I. We also determined the effect of incubation of fibroblasts with
Hp
to check its influence on fibroblast proliferation by assessing Ki67, a marker
of cell proliferation. An attempt was made to examine the effect of
Hp
on induction of HSP70 in cultured rat gastric fibroblasts and expression of
proapoptotic protein Bax.
MATERIALS AND METHODS
Cell isolation technique and fibroblast infection with Helicobater pylori
Gastric samples were harvested from 8-week-old Spraque-Dowley rats and extensively
washed with sterile phosphate-buffered saline (PBS) to remove contaminating
debris. Primary fibroblast culture was established by mincing gastric biopsy
into 12 mm
3 pieces with scissors and placing
it in tissue culture flasks under sterile conditions. Growth medium RPMI-10
containing 10% fetal bovine serum and antibiotics were added and gently mixed
with minced tissue. The flasks were maintained in a humidified atmosphere of
5% CO
2 at 37°C, and the medium was changed every
2 days. When the cells grew up to 80% of confluence and they were passaged using
standard trypsinization techniques to establish a secondary cell culture as
reported before (32, 33).
Helicobacter pylori culture
Stock cultures were maintained at 70°C in Brucella Broth (Becton Dickinson, Franklin Lakes, USA) supplemented with 10% fetal bovine serum and 10% glycerol. The
Hp strain expressing CagA and Vac cytotoxins (ATCC 700824, cagA+ vacA+) bacteria was purchased from American Type Culture Collection, and grown on Columbia Agar supplemented with 5% fresh horse blood (BioMerieux, Marcy lEtoile, France). The culture plates were incubated under microaerophilic conditions at 37°C for 35 days. Before the coincubation with fibroblasts,
Hp strain was suspended in sterile PBS and transferred to the dishes containing fibroblasts.
The sub-confluent fibroblasts were infected with 1x10
9
of live
Hp per dish and incubated in humidified atmosphere for 3, 24
and 48 hours. After incubation period, the total cellular RNA was isolated according
to procedure reported before (34).
Expression of -smooth
muscle actin, collagen I, heat shock protein-70, heme oxygenase-1, Bax, Ki67
and hypoxia inducible factor-1
transcripts in the rat gastric fibroblasts determined by real time-PCR
Expression of
-SMA,
collagen I, HSP70, HO-1, Bax, Ki67 and HIF-1
transcripts in the rat gastric fibroblasts was determined by RT-PCR, using specific
primers (
Table 1). Briefly, after the subsequent time of
Hp incubation
with fibroblasts, the cells were harvested and total cellular RNA was isolated
according to Chomczynski and Sacchi method (34) using Trizol Reagent (Invitrogen,
Carlsbad, USA) according to the manufacturers protocol. First strand cDNA was
synthesized from total cellular RNA (2 µg) using Reverse Transcription System
(Promega, Madison, USA). The PCR was carried out in an automatic DNA thermal
cycler, using 1 µg cDNA and Promega PCR reagents. For amplification of
-SMA,
collagen I, HSP70, HO-1, Bax, Ki67 and HIF-1
cDNA, gene-specific primers were used (SIGMA-Aldrich St. Louis, USA) (
Table
1). Amplification of control rat ß-actin was performed on the same
samples to verify the RNA integrity. PCR products were separated by electrophoresis
in 2% agarose gel containing 0.5 µg/ml ethidium bromide and then visualized
under UV light. Location of predicted PCR product was confirmed by using OGene
Ruler 50 bp DNA ladder (Fermentas, Life Sciences, San Francisco, USA) as a standard
marker.
| Table 1. Rat oligonucleotide
primers for detection of mRNA by RT-PCR, annealing temperature and size
of PCR products employed in the experimental protocol. |
 |
Statistical analysis
Statistical analysis of the data were performed with the use of Excel. Results
are expressed as means ±S.E.M. from 6 samples per each group. Statistical significance
of difference was determined using analysis of variance one-way ANOVA test.
Further statistical analysis for
post hoc comparisons was carried out
with Scheffe test. Differences were considered as statistically significant
at p<0.05.
RESULTS
Primary and secondary rat fibroblast culture
After harvesting gastric biopsy samples the primary fibroblast culture was established
as shown in
Fig. 1A. To confirm purity of fibroblasts in cell culture
and to avoid contamination with epithelial cells, the primary culture was passaged
three times and cultured in RPMI-1640 medium (
Fig. 1B).
 |
| Fig.
1. (A) Primary rat gastric fibroblast culture. The picture
shows fibroblasts leaving the the tissue explant one week after culture
establishement. (B) Secondary rat gastric fibroblast culture. The
secondary culture was obtained from primary culture by trypsynisation
and placing cells into separate culture dish. |
Differentiation of fibroblasts exposed to Helicobacter pylori
Differentiation of fibroblasts into myofibroblasts was assessed by determination
of expression of myofibroblast marker
-SMA
and by assessing changes in collagen I production. Using specific primers for
-SMA, collagen
I and ß-actin as an internal standard, single bands for each of cDNAs
at the expected size were observed. The ratio of
-SMA
and collagen I to ß-actin for intact secondary fibroblast culture (control)
and for 3, 24 and 48 hours of incubation with
Hp are shown in
Figs.
2A and
3A. Densitometric analysis of mRNAs expression in control
(not exposed to
Hp) cells showed moderate level of
-SMA
and relatively low level of collagen I and these effects were not significantly
influenced by coincubation of fibroblasts with
Hp at 3 h and 24 h, respectively
(
Figs. 2B and
3B). In contrast, ratio of mRNA expression (
-SMA
and ß-actin) level revealed that the expression of
-SMA
mRNA was significantly upregulated at 48 h of co-culture with
Hp. The
parallel increase in expression of collagen I mRNA was recorded in fibroblasts
co-cultured with
Hp at 48 h following bacteria application (
Figs.
2B and
3B).
 |
Fig. 2. RT-PCR analysis of
mRNA expression for -SMA
(panel A) in rat gastric fibroblasts at 3, 24 and 48 h of co-incubation
with Hp strain CagA+, VacA+ (ATTC 700824) and the ratio of -SMA
over ß-actin (panel B) at respective times of co-incubation
with this bacteria. Results are mean ±S.E.M. of 6 determinations.
Asterisk indicates a significant (p<0.05) change as compared to the
value obtained at 3 h and 24 h of incubation with Hp. |
 |
Fig. 3. RT-PCR analysis of
mRNA expression for collagen I (panel A) in rat gastric fibroblast
at 3, 24 and 48 h of co-incubation with Hp (ATTC 700824) and the
ratio of collagen I over ß-actin (panel B) at respective times
of co-incubation of these cells with bacteria. Results are mean ±S.E.M.
of 6 determinations. Asterisk indicates a significant change (p<0.05)
above these recorded at 3 h and 24 h of co-incubation with Hp. |
Expression of heat shock protein-70 and heme oxygenase-1 mRNA, the apoptosis related gene Bax and proliferation marker Ki67 in cultured fibroblasts with or without
co-incubation with Helicobater pylori
The mRNA for stress and pro-apoptotic proteins were assessed in culture of gastric
fibroblasts. As shown in
Figs. 4A and
4B, the incubation of cagA
and vacA positive
Hp strain with gastric fibroblasts upregulated in a
time-dependent manner the expression of HSP70 mRNA in these cells. This significant
increase in HSP70 mRNA expression was observed already after 24 hours and subsequently
noted at 48 h after exposure of cell culture to this bacteria. In contrast,
the cagA and vacA positive
Hp strain had no significant influence on
the expression of HO-1 mRNA level in fibroblasts in any of the selected times
of the incubation (from 3 h to 48 h) (
Figs. 5A and
5B).
 |
Fig. 4. RT-PCR analysis of
mRNA expression for HSP70 (panel A) in rat gastric fibroblast at
3, 24 and 48 h of co-incubation with Hp (ATTC 700824) and the ratio
of HSP70 over ß-actin (panel B) at respective times of co-incubation
of these cells with Hp. Results are mean ±S.E.M. of 6 determinations.
Asterisk indicates a significant change (p<0.05) as compared to the
value obtained at 3 h of incubation with Hp. |
 |
Fig. 5. RT-PCR analysis of
mRNA expression for HO-1 (panel A) in rat gastric fibroblast at
3, 24 and 48 h of co-incubation with Hp (ATTC 700824) and the ratio
of HO-1 over ß-actin (panel B) at respective times of co-incubation
of these cells with Hp. Results are mean ±S.E.M. of 6 determinations. |
Next we examined the expression of mRNA for pro-apoptotic Bax and the expression
of mRNA for proliferation marker Ki67 in fibroblasts exposed to
Hp strain
expressing CagA and VacA cytotoxins. As shown in
Figs. 6A and
6B
the expression of Bax mRNA was significantly decreased already after 3 hours
of co-incubation with this bacteria and this significant inhibition persisted
throughout 24 h and 48 h upon start of incubation. At 48 h of incubation a significant
inhibition of the signal for Bax mRNA was observed below the value of Bax mRNA
recorded at 3 h and 24 h. The signal of mRNA for Ki67 was also unchanged at
3 h of incubation of fibroblasts with
Hp expressing CagA and VacA cytotoxins
compared to vehicle control (
Figs. 7A and
7B).
 |
Fig. 6. RT-PCR analysis of
mRNA expression for Bax (panel A) in rat gastric fibroblast at
3, 24 and 48 h of co-incubation with Hp (ATTC 700824) and the ratio
of Bax over ß-actin (panel B) at respective times of co-incubation
of these cells with Hp. Results are mean ±S.E.M. of 6 determinations.
Asterisk indicates a significant change (p<0.05) as compared to the
value obtained in control cells. Asterisk and cross indicate a significant
change (p<0.05) as compared to the value obtained at 24 h. |
 |
Fig. 7. RT-PCR analysis of
mRNA expression for Ki67 (panel A) in rat gastric fibroblast at
3, 24 and 48 h of co-incubation with Hp (ATTC 700824) and the ratio
of Ki67 over ß-actin (panel B) at respective times of co-incubation
of these cells with Hp. Results are mean ±S.E.M. of 6 determinations. |
At 24 h of fibroblasts incubation with this bacteria, the signal for Ki67 in
fibroblasts failed to show significant difference compared with that recorded
at 3 h of this incubation. Also proliferation at 48 h of coincubation with
Hp
remained unchanged as it was estimated by proliferation marker Ki67 level of
mRNA (
Figs. 7A and
7B).
Determination of hypoxia inducible factor-1
mRNA level in fibroblasts co-incubated with Helicobacter pylori
The signal of mRNA for HIF-1
was not significantly altered at 3 h of incubation of fibroblasts with alive
Hp CagA+, VacA+ (
Figs. 8A and
8B). However, the mRNA for
HIF-1
was significantly
increased (p<0.05) over that measured at 3 h. This significant upregulation
of HIF-1
gene had
persisted at 48 h upon the incubation with this bacteria and the ratio of HIF-1
over ß-actin mRNA reached significantly higher value over that determined
at 24 h of incubation. Increase in HIF-1
mRNA expression was highly significant after 48 hours of fibroblast co-culture
with
Hp (
Figs. 8A and
8B) parallel with the increase in
expression of fibroblasts differentiation marker
-SMA
(
Fig. 1B).
 |
Fig. 8. RT-PCR analysis of
mRNA expression for HIF-1
(panel A) in rat gastric fibroblast at 3, 24 and 48 h of co-incubation
with Hp (ATTC 700824) and the ratio of HIF-1
over ß-actin (panel A) at respective times of co-incubation of
these cells with Hp (panel B). Asterisk indicates a significant
change (p<0.05) compared to the value obtained at 3 h of incubation
with Hp. Results are mean ±S.E.M. of 6 determinations. Asterisk
indicates a significant change (p<0.05) as compared to the value obtained
in control cells. |
DISCUSSION
The involvement of
Hp in the pathogenesis of a variety of gastric disorders, including peptic ulcers, MALT lymphoma and gastric cancer is well documented, however, the importance of other non-epithelial cellular compartments in
Hp pathology of the gut has been little studied. Impaired balance between aggresive bacterial and host defensive factors plays an essential role in the different clinical outcomes of
Hp infection in the host. As the stromal microenvironment is a complex structure composed of an extracellular matrix (ECM), activated fibroblasts and myofibroblasts, inflammatory cells, blood and lymphatic vessels, there is convincing evidence that any change in that system, can influence the normal architecture of epithelial cells and alter their functions by this non-epithelial component.
Therefore, the aim of our present study was to determine, whether gastric fibroblasts may represent cellular target of
Hp affecting process of proliferation, apoptosis and cell defense system. We assumed that the increased differentiation into myofibroblasts, collagen production and changes in proliferation and apoptosis rates can be implicated in the mechanism of interaction between fibroblasts and
Hp. Our data presented in this work indicate that
Hp increased differentiation of fibroblasts into myofibroblasts because the expression of
-SMA was enhanced by this bacteria. This also indicates that stroma cells could be affected by
Hp suggesting that fibroblasts could act as the one of the major source of myofibroblasts.
Moreover, we found that the enhanced production of
-SMA correlated with the expression of mRNA for collagen type 1. That is in agreement with existing evidence showing that myofibroblasts positive for
-SMA correlate with enhanced production and deposition of extracellular matrix related proteins including collagen type I - an event associated with fibrosis (15, 18, 19, 35, 36). Moreover, the presence of abundant myofibroblasts in gastric carcinoid tumours has been postulated (20).
Interactions between the neoplastic and non-neoplastic cells and extracellular
matrix have been proposed to stimulate the extensive desmoplastic reaction (19,
35). At the molecular level, stroma production is promoted by the activation
of multiple cancer cell-derived signaling pathways such as transforming growth
factor ß (TGF-ß), hepatocyte growth factor (HGF/Met), fibroblast
growth factors (FGFs), insulin-like growth factor 1 (IGF-1) and epidermal growth
factor (EGF) acting
via autocrine and paracrine mechanisms (19, 35).
These receptor-mediated signaling cascades activated by growth factors lead
to secretion of structural matrix components including proteoglycans, collagens
and fibronectin as well as catalytically active enzymes such as proteinases
(35).
Generally it is accepted, that when the epithelium changes, the stroma inevitably
follows. In cancer, changes in the stroma drive invasion and metastasis, the
hallmarks of malignancy. As it was mentioned before stromal changes at the invasion
front include the appearance of myofibroblasts that take a part in various aspects
of the cancer progression increased cell invasion, and the suppression of apoptosis
(19, 35). The stimulation of stromal cells enhances the production of growth
factors such as HGF in turn acting on epithelial cells (35). For instance,
-SMA
next to PDGFB-R is in fact regarded as CAFs (cancer-associated fibroblasts)
marker (36). According to literature the transdifferentiation of fibroblasts
into myofibroblasts is mediated by a cancer cell-derived cytokines such as TGF-ß
(15). Here we show, that the presence of
Hp and direct contact of
Hp
with fibroblasts itself can induce this process directly and it is capable of
influencing the stroma architecture, without prior interactions with epithelial
cells.
The concept suggesting that the microenvironment is crucial for the mainteinance of cellular functions and tissue integrity indicates that a neoplasia-induced change in the stroma may contribute to cancer invasion (37-39). Experimental animal models have demonstrated that cancer invasion is stimulated by the wound-healing stroma (37-39). Therefore, in both wound healing and tumorgenesis, the transition of fibroblasts to myofibroblasts is an example of the stromal alteration. The casual role of myofibroblasts in the transition from the noninvasive towards the invasive phenotype is supported by the evidence that the appearance of myofibroblasts precedes the invasive stage of cancer (37-39). Experimental and clinical observations seem to suggest that myofibroblasts may be proinvasive and capable of producing proinvasive signals (37-39).
The evidence based scientific observations suggest that growth factors such
as TGF-ß1 implicated in wound healing (38, 39) may also play a role in
changing of the stromal host compartment in cancer. TGF-ß is known to
upregulate
-SMA
actin expression in fibrocytes and transdifferentiate them into myofibroblasts
in many systems (15, 21, 37-39).
We also observed the enhancement in expression of mRNA for TGF-ß (data
not shown). But as given literature evidence, TGF-ß is secreted as a latent
complex that requires an activation that is regulated environmentally, either
proteolitically or nonproteolitically. Host cells such myofibroblasts may play
important role in turning TGF-ß into biologically active, since they are
considered as major suppliers of proteinases and trombospondin (41). Fibroblast
to myofibroblast transdifferentiation may be responsible for an activation of
TGF-ß in tumours (38-41).
The increase in myofibroblasts in colon cancer could result from transdifferentiation of resident interstitial fibroblasts into myofibroblasts or a decreased rate of apoptosis (38, 41). Our results suggest both transdifferentiation and decreased apoptosis in
Hp infected fibroblasts.
The quantity of ECM proteins, provide structural and functional scaffolding
for cancer cells
i.e. cytoskeletal organization, ectopic survival and
differentiation (41) as well as shielding cancer cells from the immune response
(41, 42). The collagen type I secretion downregulates E-cadherin gene expression
and stabilizes epithelial to myofibroblast transdifferentiation (38, 41, 42).
Since myofibroblasts are potentially invasive, they may invade collagen I, so
they may be able to migrate, invade and stimulate cancer cells and
vice versa.
We found the enhanced upregulation of collagen I which is in agreement with the studies suggesting that CagA-positive strain of
Hp plays an important role in tissue remodeling, angiogenesis, cancer invasion and metastasis (43, 45). Moerover, studies in whole blood cultures revealed that
Hp strain expressing CagA cytotoxin exhibited immunomodulatory effect on expression of CD25 and CD58 receptors on lymphocytes and activation of cytokines such as IL-10 capable of inhibit the proimflammatory activity of monocytes and T lymphocytes (45). We propose, that some of these effects may be mediated by myofibroblasts because after eradication of
Hp inflammatory cell infiltration is reduced, but the advanced intestinal metaplasia mucosa itself is usually not improved (46). The accumulation and activation of HIF-1
is a very challenging field of study since the responses are cell type specific and, therefore, may vary between cells of different origin. There is an increasing body of evidence that HIF-1
plays an important role in various infections (26, 30, 43, 44), however, the mechanisms of HIF-1
activation by
Hp are not well established. The study of Bhattacharyya
et al. (27) have indicated that
Hp-mediated APE1 induction enhanced HIF-1
expression and, along with p300, upregulated HIF-1
transcriptional activity in the gastric epithelium. A cell line study has shown, that the reactive oxygen species produced by
Hp, stabilise HIF-1
, leading to its increased expression (28). The similar effect has been observed by Ding and coworkers (47).
The
Hp expressing CagA cytotoxin and also CagA by itself can induce ROS
production
via NADPH oxidase-dependent pathway (48) causing DNA damage,
thus contributing to apoptosis in early stages of gastric carcinogenesis. However,
this was not the case with respect to apoptosis in our study because Bax expression
was downregulated in fibroblasts incubated with
Hp. In culture of MCF7
cancer cells and fibroblasts, the enhanced ROS production and oxidative stress
in adjacent fibroblasts as well as the activation of HIF-1
-
and NF
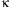
B driven
gene transcription were observed. In their study (49), ROS were responsible
for activation of myofibroblast markers and extracellular matrix proteins associated
with activated fibroblasts. Therefore, we attempted to check if the same effect
appears in our system, especially that, according to existing evidence, many
of the proteins induced by activated HIF-1
in fibroblasts are highly expressed in the tumor stroma of human breast cancer
patients being particularly responsible for tumor recurrence or metastasis (49,
50). HIF-1
acts
as a tumor promoter in stromal cells but as a tumor suppressor in cancer cells.
Tumour hypoxia is now recognised as a key factor driving the development of malignancy, and the master regulatory protein in the response of cells to change in oxygen levels
via HIF-1
(49, 50). We postulate here that HIF-1
could represent one of the prerequisites to development of gastric cancer development in
Hp infected mucosa. This is supported by the fact that HIF-1
expression was upregulated in fibroblast co-incubated with
Hp.
As there is increasing evidence that apoptosis plays an important role in the pathogenesis of a variety of infectious diseases, we decided to check whether incubation of gastric fibroblasts with
Hp influences the expression of HSP70, apoptosis related proteins Bax and expression of proliferation marker Ki67. In our culture model system, the cagA and vacA positive
Hp strain upregulated in a time-dependent manner the expression of HSP70 mRNA in fibroblasts. This increase in HSP70 mRNA expression was observed already after 24 hours and subsequently noted at 48 h after exposure of cell culture to
Hp. The expression of Bax mRNA significantly decreased in fibroblasts already after 3 hours of their co-incubation with bacteria. This finaling indicates that
Hp inhibits apoptosis of fibroblasts
in vitro.
Previous studies revealed that epithelial cells co-incubation with
Hp
induces apoptosis and causes a fall in expression of HSP70 (22). This excessive
apoptosis resulted in tissue damage. Targosz
et al. (22) found that
cagA,
vacA positive
Hp strains enhanced apoptosis in rat gastric cancer
epithelial cells, as evidenced by an increase in expression of pro-apoptotic
Bax and decrease in expression for anti-apoptotic Bcl-2. At present, the mechanism
by which
Hp induces apoptosis in gastric cancer cells and interacts with
HSP70 expression remains unknown. The major finding of their study (22) was
that
Hp can directly attenuate the expression of HSP70 in MKN7 cells
which could accompany the rise in apoptosis. It has been postulated that inactivation
of HSP90 and HSP70 leads to loss of invasion in a variety of cancer cell types
(51). Thus, we conclude that the influence of
Hp on HSP70 may be cell
type specific. In the case of epithelial cells,
Hp evoked an increase
in apoptosis, while in the case of fibroblasts this bug evidently promoted their
differentiation with sustained rate of proliferation.
It has been demonstrated that HO-1 mRNA and protein levels are strongly inducible in fibroblasts following i.e. oxidative stress (UVA radiation and hydrogen peroxide) (52, 53). HO-1 activation exhibits a cytoprotective effect due to the anti-inflammatory, anti-apoptotic activity, and preserved control in regulation of proliferative activity (25, 26). In our experimental model the HO-1 mRNA was also strongly expressed, but we observd no alteration in HO-1 expression level following co-incubation with
Hp. Lakkisto
et al. (53) showed that, HO-1 decreased accumulation and proliferation of fibroblasts, and down-regulated procollagen type I expression in the infarct area of heart failure model in rats. Lack of major alteration in HO-1 expression under our experimental settings is in agreement with the studies pointing to antifibrogenic properties of HO-1. According to their accumulated evidence (53) the cardioprotective effects of HO-1 in the late phase of infarct healing may be mediated by the down-regulation of the profibrotic connective tissue growth factor (CTGF), because HO-1 decreased CTGF expression at week 4 in their study (53).
Nakamura
et al. (54) showed that increased induction of HO-1 exerts suppressive effects on the expression of collagen and this also counteracted changes in EMT gene changes. This depend upon a specific increase in HO-1 expression caused by cobalt protoporphyrin prevented CsA-mediated
-SMA induction, whereas genetic inhibition of HO-1 by siRNA substantially enhanced
-SMA induction compared to control cells (55). The enhanced differentiation and fibrinogenesis caused by upregulation od HO-1 expression depends on the cell specificity (56).
Taken together, the presence of
Hp had no effect on the expression of HO-1 mRNA in our fibroblasts system. Our cells were not stimulated prior to infection with
Hp which is consistent with the recent observation that
Hp inhibits NO-stimulated HO-1 expression in gastric epithelial cells through a mechanism that requires CagA (57). Our results seem to be also consistent with the statement that
Hp favors its own pathogenesis by limiting HO-1 induction (57), stimulation of fibroblast differentiation, sustaining of proliferation and matrix deposition. Another way to confirm these findings, it will be necessary to stimulate cells before subsequent infection with
Hp.
We conclude, that the exposure of fibroblasts to
Hp promotes their differentiation into myofibroblasts, with possible excess of matrix production
in vitro. The increased expression of HSP70 mRNA suggests that induction of HSP70 confers cytoprotection against
Hp infection, by inhibiting the expression of Bax. Fibroblasts seem to cooperate with epithelial component in the stomach infected with
Hp, that may facilitate inflammation and peptic ulcer formation with potential risk to develop gastric cancer. However, further studies in fibroblasts affected by
Hp and analysis of factors influenced by this bug at the protein level as well as the effect of incubation with various strains of
Hp on the complex fibroblast-epithelial cell interaction are warranted.
Acknowledgements:
Supported by Jagiellonian University Medical College grant: K/ZDS/003042 to
Drs. G. Krzysiek-Maczka and A. Targosz
Conflict of interests: None declared.
REFERENCES
- Konturek PC, Konturek SJ, Brzozowski T. Helicobacter pylori infection
in gastric cancerogenesis. J Physiol Pharmacol 2009; 60: 3-21.
- Blaser MJ. Heterogeneity of Helicobacter pylori. Eur J Gastroenterol
Hepatol 2012; 9(Suppl. 1): S3-S6.
- Asaka M, Kato M, Graham DY. Prevention of gastric cancer by Helicobacter
pylori eradication. Intern Med 2010; 49: 633-636.
- Suzuki M, Kiga K, Kersulyte D, et al. Attenuated CagA oncoprotein
in Helicobacter pylori from Amerindians in Peruvian Amazon.
J Biol Chem 2011; 286: 29964-29972.
- Uemura N, Okamoto S, Yamamoto S, et al. Helicobacter pylori
infection and the development of gastric cancer. N Engl J Med 2001; 345:
784-789.
- Amieva MR, El-Omar EM. Host-bacterial interactions in Helicobacter
pylori infection Gastroenterology 2008; 134: 306-323.
- Crabtree JE. Immune and inflammatory responses to Helicobacter pylori
infection. Scand J Gastroenterol 1996; 215: 3-10.
- Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, Haas R. Translocation
of Helicobacter pylori CagA into gastric epithelial cells by type
IV secretion. Science 2000; 287: 1497-1500.
- Stein M, Bagnoli F, Halenbeck R, Rappuoli R, Fantl WJ, Covacci A. c-Src/Lyn
kinases activate Helicobacter pylori CagA through tyrosine phosphorylation
of the EPIYA motifs. Mol Microbiol 2002; 43: 971-980.
- Birchmeier C, Birchmeier W, Gherardi E, Vande Woude GF. Met, metastasis,
motility and more. Nat Rev Mol Cell Biol 2003; 4: 915-925.
- Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat
Rev Cancer 2002; 2: 442-454.
- Scotti C, Mignosi P, Filipazzi P, et al. Molecular alterations
in fibroblasts exposed to Helicobacter pylori broth culture filtrate:
a potential trigger of autoimmunity? Helicobacter 2010; 15: 76-77.
- Hemers E, Duval C, McCaig C, Handley M, Dockray GJ, Varro A. Insulin-like
growth factor binding protein-5 is a target of matrix metalloproteinase-7:
implications for epithelial-mesenchymal signaling. Cancer Res 2005; 65:
7363-7369.
- McCaig C, Duval C, Hemers E, et al. The role of matrix metalloproteinase
(MMP)-7 in redefining the gastric microenvironment in response to Helicobacter
pylori. Gastroenterology 2006; 130: 1754-1763.
- Lin A, Hokugo A, Nishimura I. Wound closure and wound management: a new
therapeutic molecular target. Cell Adh Migr 2010; 4: 396-399.
- Vandekerckhove J, Weber K. Actin typing on total cellular extracts: a
highly sensitive protein-chemical procedure able to distinguish different
actins. Eur J Biochem 1981; 113: 595-603.
- Gabbiani G. Modulation of fibroblastic cytoskeletal features during wound
healing and fibrosis. Pathol Res Pract 1994; 190: 851-853.
- Duffield JS, Lupher M, Thannickal VJ, Wynn TA. Host responses in tissue
repair and fibrosis. Annu Rev Pathol 2013; 8: 241-276.
- Koyama S. Coordinate cell-surface expression of matrix metalloproteinases
and their inhibitors on cancer-associated myofibroblasts from malignant
ascites in patients with gastric carcinoma. J Cancer Res Clin Oncol 2005;
131: 809-814.
- Ishikawa T, Ando T, Obayashi H, et al. Helicobacter pylori
isolated from a patient with Menetriers disease increases hepatocyte growth
factor mRNA expression in gastric fibroblasts: comparison with Helicobacter
pylori isolated from other gastric diseases. Dig Dis Sci 2008; 53: 1785-1791.
- Trujillo KA, Heaphy CM, Mai M, et al. Markers of fibrosis and epithelial
to mesenchymal transition demonstrate field cancerization in histologically
normal tissue adjacent to breast tumors. Int J Cancer 2011; 129: 1310-1321.
- Targosz A, Brzozowski T, Pierzchalski P, et al. Helicobacter
pylori promotes apoptosis, activates cyclooxygenase (COX)-2 and inhibits
heat shock protein HSP70 in gastric cancer epithelial cells. Inflamm Res
2012; 61: 955-966.
- Liu ZM, Chen GG, Ng EK, Leung WK, Sung JJ, Chung SC. Upregulation of heme
oxygenase-1 and p21 confers resistance to apoptosis in human gastric cancer
cells. Oncogene 2004; 23: 503-513.
- Yin Y, Liu Q, Wang B, Chen G, Xu L, Zhou H. Expression and function of
heme oxygenase-1 in human gastric cancer. Exp Biol Med (Maywood) 2012; 237:
362-371.
- Was H, Dulak J, Jozkowicz A. Heme oxygenase-1 in tumor biology and therapy.
Curr Drug Targets 2010; 11: 1551-1570.
- Wagener FA, Scharstuhl A, Tyrrell RM, et al. The heme-heme oxygenase
system in wound healing; implications for scar formation. Curr Drug Targets
2010; 11: 1571-1585.
- Bhattacharyya A, Chattopadhyay R, Hall E, Mebrahtu S, Ernst P, Crowe S.
Mechanism of hypoxia-inducible factor 1a-mediated Mcl1 regulation in Helicobacter
pylori-infected human gastric epithelium. Am J Physiol Gastrointest
Liver Physiol 2010; 299: G1177-G1186.
- Park JH, Kim TY, Jong HS, et al. Gastric epithelial reactive oxygen
species prevent normoxic degradation of hypoxia-inducible factor-1alpha
in gastric cancer cells. Clin Cancer Res 2003; 9: 433-440.
- Zhong H, De Marzo AM, Laughner E, et al. Overexpression of hypoxia-inducible
factor 1alpha in common human cancers and their metastases. Cancer Res 1999;
59: 5830-5835.
- Chiavarina B, Whitaker-Menezes D, Migneco G, et al. HIF1-alpha
functions as a tumor promoter in cancer associated fibroblasts, and as a
tumor suppressor in breast cancer cells: autophagy drives compartment-specific
oncogenesis. Cell Cycle 2010; 9: 3534-3551.
- Whitaker-Menezes D, Martinez-Outschoorn UE, Lin Z, et al. Evidence
for a stromal-epithelial lactate shuttle in human tumors: MCT4 is a marker
of oxidative stress in cancer-associated fibroblasts. Cell Cycle 2011; 10:
1772-1783.
- Krzysiek-Maczka G, Michalik M, Madeja Z, Korohoda W. Involvement of cytoskeleton
in orientation of cell division in contact guided cells. Folia Biol (Krakow)
2010; 58: 21-27.
- Krzysiek-Maczka G, Korohoda W. Surface anisotropy orients cell divisions
in contact guided cells. Folia Biol (Krakow) 2008; 56: 13-19.
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium
thiocyanate-phenol-chloroform extraction. Anal Biochem 1987; 162: 156-159.
- Varro A, Kenny S, Hemers E, et al. Increased gastric expression
of MMP-7 in hypergastrinemia and significance for epithelial-mesenchymal
signaling. Am J Physiol Gastrointest Liver Physiol 2007; 292: G1133-G1140.
- Kumar S, Weaver VM. Mechanics, malignancy, and metastasis: the force journey
of a tumor cell. Cancer Metastasis Rev 2009; 28: 113-127.
- De Wever O, Westbroek W, Verloes A, et al. Critical role of N-cadherin
in myofibroblast invasion and migration in vitro stimulated by colon-cancer-cell-derived
TGF-beta or wounding. J Cell Sci 2004; 117: 4691-4703.
- De Wever O, Mareel M. Role of tissue stroma in cancer cell invasion. J
Pathol 2003; 200: 429-447.
- De Wever O, Nguyen QD, van Hoorde L, et al. Tenascin-C and SF/HGF
produced by myofibroblasts in vitro provide convergent pro-invasive
signals to humancolon cancer cells through RhoA and Rac. FASEB J 2004; 18:
1016-1018.
- De Wever O, Demetter P, Mareel M, Bracke M. Stromal myofibroblasts are
drivers of invasive cancer growth. Int J Cancer 2008; 123: 2229-2238.
- Nazareth MR, Broderick L, Simpson-Abelson MR, Kelleher RJ, Yokota SJ,
Bankert RB. Characterization of human lung tumor-associated fibroblasts
and their ability to modulate the activation of tumorassociated T cells.
J Immunol 2007; 178: 5552-5562.
- Iwamoto J, Mizokami Y, Takahashi K, et al. Expressions of urokinase-type
plasminogen activator, its receptor and plasminogen activator inhibitor-1
in gastric cancer cells and effects of Helicobacter pylori. Scand
J Gastroenterol 2005; 40: 783-793.
- Tegtmeyer N, Wittelsberger R, Hartig R, Wessler S, Martinez-Quiles N,
Backert S. Serine phosphorylation of cortactin controls focal adhesion kinase
activity and cell scattering induced by Helicobacter pylori. Cell
Host Microbe 2011; 9: 520-531.
- Mutoh H, Sashikawa M, Hayakawa H, Sugano K. Monocyte chemoattractant protein-1
is generated via TGF-beta by myofibroblasts in gastric intestinal
metaplasia and carcinoma without H. pylori infection. Cancer Sci 2010; 101:
1783-1789.
- Wiese M, Eljaszewicz A, Andryszczyk M, et al. Immunomodulatory
effects of Lactobacillous plantarum and Helicobacter pylori CagA+
on the expression of selected superficial molecules on monocyte and lymphocyte
and the synthesis of cytokines in whole blood culture. J Physiol Pharmacol
2012; 63: 217-224.
- Ding SZ, Minohara Y, Fan XJ, et al. Helicobacter pylori
infection induces oxidative stress and programmed cell death in human gastric
epithelial cells. Infect Immun 2007; 75: 4030-4039.
- Martinez-Outschoorn UE, Balliet RM, Rivadeneira DB, Chiavarina B, Pavlides
S, Wang C. Oxidative stress in cancer associated fibroblasts drives tumor-stroma
co-evolution: a new paradigm for understanding tumor metabolism, the field
effect and genomic instability in cancer cells. Cell Cycle 2010; 9: 3256-3276.
- Cha B, Lim JW, Kim KH, Kim H. 15-Deoxy-D12,14-prostaglandin J2 suppresses
RANTES expression by inhibiting NADPH oxidase activation in Helicobacter
pylori-infected gastric epithelial cells. J Physiol Pharmacol 2011;
62: 167-174.
- Chiavarina B, Whitaker-Menezes D, Migneco G, et al. HIF-1 alpha
functions as a tumor promoter in cancer associated fibroblasts, and as a
tumor suppressor in breast cancer cells: autophagy drives compartment-specific
oncogenesis. Cell Cycle 2010; 9: 3534-3551.
- Sims JD, McCready J, Jay DG. Extracellular heat shock protein Hsp70 and
Hsp90a assist in matrix metalloproteinase-2 activation and breast cancer
cell migration and invasion. PLoS One 2011; 6: e18848.
- Applegate LA, Noel A, Vile G, Frenk E, Tyrrell RM. Two genes contribute
to different extents to the heme oxygenase enzyme activity measured in cultured
human skin fibroblasts and keratinocytes: implications for protection against
oxidant stress. Photochem Photobiol 1995; 61: 285-291.
- Zhong JL, Edwards GP, Raval C, Li H, Tyrrell RM. The role of Nrf2 in ultraviolet
A mediated heme oxygenase 1 induction in human skin fibroblasts. Photochem
Photobiol Sci 2010; 9: 18-24.
- Lakkisto P, Siren JM, Kyto V, et al. Heme oxygenase-1 induction
protects the heart and modulates cellular and extracellular remodelling
after myocardial infarction in rats. Exp Biol Med (Maywood) 2011; 236: 1437-1448.
- Nakamura T, Matsushima M, Hayashi Y, et al. Attenuation of transforming
growth factor-ß-stimulated collagen production in fibroblasts by quercetin-induced
heme oxygenase-1. Am J Respir Cell Mol Biol 2011; 44: 614-620.
- Shin DH, Park HM, Jung KA, et al. The NRF2-heme oxygenase-1 system
modulates cyclosporin A-induced epithelial-mesenchymal transition and renal
fibrosis. Free Radic Biol Med 2010; 48: 1051-1063.
- Yang H, Zhao LF, Zhao ZF, Wang Y, Zhao JJ, Zhang L. Heme oxygenase-1 prevents
liver fibrosis in rats by regulating the expression of PPAR
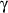 and NF-B.
World J Gastroenterol 2012; 18: 1680-1688.
and NF-B.
World J Gastroenterol 2012; 18: 1680-1688.
- Gobert AP, Asim M, Piazuelo MB, et al. Disruption of nitric oxide
signaling by Helicobacter pylori results in enhanced inflammation
by inhibition of heme oxygenase-1. J Immunol 2011; 187: 5370-5379.