CITICOLINE IMPROVED CARDIOVASCULAR FUNCTION
IN ANIMAL MODEL OF DYSAUTONOMIA
INTRODUCTION
The integrity of the cardiac functions depends on the optimal heart-nervous system connections. The heart is innervated by the autonomic nervous system, classified into sympathetic and parasympathetic nervous systems. Despite the opposite functions of the two systems, to maintain homeostasis, sympathetic and parasympathetic systems play an optimal role in regulating cardiovascular functions (1). Parasympathetic cardiac innervation provides an inhibitory impact by slowing down the heart rate , the neural control of the arterial blood pressure is the outcome of the activities of both systems on the central and peripheral levels (2). In contrast, sympathetic innervation accelerates the ventricular contraction and increases the heart rate by influencing neurotransmitter norepinephrine.
Loss of catecholaminergic innervation of the cardiac tissues results in sympathetic neurodegeneration of the heart and is considered a cardiac dysautonomia mechanism (3). Interestingly, patients with Parkinson’s disease exhibit varying levels of cardiac denervation. However, in Parkinson’s disease patients with intimately related cardiac dysautonomia and neurodegeneration, various symptoms characterize each component. Cardiac dysautonomia in Parkinson’s disease refers to cardiac neurodegeneration, orthostatic hypotension, fatigue, decreased time to peak heart rate variability, shortness of breath at regular exercise, and reduced blood catecholamines, especially plasma norepinephrine (4, 5).
Several animal models of cardiac sympathetic dysautonomia and neurodegeneration are available to study and validate therapeutic targets for preclinical evaluation. MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) is a lipophilic neurotoxin widely used in Parkinson’s disease nigral dopaminergic loss in mice (6) and monkeys (7). It has the ability not only to cross the blood-brain barriers affecting brain stem nuclie the sympathetic autonomic nuclei such as rostroventrolateral medulla (RVLM) and nucleus tractus solitarii (NTS) in the lower hindbrain, causing desensitization of baroreceptor reflex and impaired heart rate variability (8) but also induces loss of cardiac sympathetic innervation (9). MPTP affects the heart and gut through catecholamine neuron degeneration and extends to chromaffin cells of the adrenal medulla (10). Loss of cardiac sympathetic nerve induced by MPTP has been previously demonstrated in rats by a daily dose (20 mg/kg/5 days, i.p.) to yield 100% loss of cardiac norepinephrine (11).
Citicoline or CDP-choline is a drug composed of a cytidine 5’-diphosphate moiety and choline, after being absorbed it is hydrolyzed into cytidine 5’-diphosphate and choline, that can cross the blood-brain barrier. In the brain, these hydrolysis derived components recombined to form citicoline in the neurons increasing the brain content of phospholipids (12). Citicoline is a safe drug with negligible acute or chronic toxicity (13).
Previous studies introduced the neuroprotective effect of cytidine-diphosphate-choline (CDP-choline or citicoline) against several neurological disorders like Alzheimer’s disease, ischemic stroke (14), spinal cord injury (15) and a recent study showed its protective effect in children after cardiac arrest (16). In the current study, we aimed to study the effects of citicoline on cardiovascular function in MPTP-treated albino rats as an animal model for cardiovascular dysautonomia.
MATERIALS AND METHODS
Animal maintenance
Twenty-four male albino rats, initially weighing 150 – 200 g, were received from the Ophthalomolgy Research Institute (Giza, Egypt). We kept the rats in conventional animal service-based housing. The rats were accommodated to normal standard conditions (temperature 24 ± 1°C, relative humidity 40 – 60%, and a 12:12 h light/dark photoperiods). Moreover, they were maintained in polycarbonated cages (3 rats/cage) to avoid isolation stress for one week before starting the experiment. The rats were kept under monitoring along the acclimatization period to exclude infectious rats. The animals were provided with a standard basal diet (El-Nasr Inc., Giza, Egypt) and water ad libitum.
The approval of animal procedures, Care, and Use of Laboratory Animals was guided based on the Ethics and Scientific Committee (Eighth edition 2011), Department of Physiology, Kasr Al Ainy, Faculty of Medicine, Cairo University, Egypt.
Experimental design for in vivo work
Pathogen-free twenty-four male albino rats were divided into four groups (6 rats each). The first group was designated as a negative control (normal control) and received intraperitoneal (i.p.) saline injection for five consecutive days. The second group was a positive control group and received citicoline (Somazina drops October Pharma, 6th of October city - Egypt Under License of Ferrer International, SA, Spain) (250 mg/kg) by oral gavage for consecutive 20 days based on a previous study (17). The third and fourth groups treated with MPTP-HCL (Sigma, St Louis, MO, USA) (30 mg/kg, i.p.) for five consecutive days based on previous studies (18), followed by additional treatment with oral doses of citicoline (250 mg/kg) for only the fourth group, which continued for 20 days. Food, water, and behavioral changes were monitored periodically, and the body weights were recorded weekly. At the end of the study, all the rats were exposed to the cardiovascular system’s functional assessment before euthanasia.
In vivo functional assessment of the cardiovascular system
The functional measurements performed over two days separated by one day (to allow recovery). D1: Transthoracic echocardiography using ketamine and xylazine, D3: measuring blood pressure in conscious rats followed ECG under urethane anesthesia.
1) Transthoracic echocardiography
The assessment was performed by using echocardiography to evaluate the cardiac functions in vivo. The rats were anesthetized by injecting both ketamine-hydrochloride (25 mg/kg, bw, i.p.) and xylazine (5 mg/kg, bw, i.p.). Anesthesia was followed by removing the hairs from the chest’s anterior part and maintaining the rats on a specialized warming table to maintain normothermia (19).
The echocardiography system was equipped with a 10 MHz phased-array transducer (GE Healthcare’s Vivid, USA). 2D short-axis view of the left ventricle and M-mode tracings were recorded to measure ejection fraction (EF), fractional shortening (FS), left ventricular dimension in diastole (LVDd), left ventricular dimension in systole (LVSd), left ventricular posterior wall thickness during diastole (LVPWD), left ventricular posterior wall thickness during systole (LVPWS), stroke volume (SV) and heart rate (HR) were estimated. Cardiac output was calculated from SV and HR [CO = HR × SV]. Trans mitral Doppler flows (ratios of E and A velocities) were recorded in an apical 4-chamber orientation with the sample volume placed at the tips of the mitral leaflet.
2) Measurement of the arterial systolic blood pressure
Blood pressure of animals were measured by non-invasive blood pressure monitor (LE 5001 , LETICA scientific Instruments, Espania) from the tail of conscious rats by the tail-cuff technique.
The tail-cuff technique is a common, non-invasive and convenient way to measure systolic blood pressure in an animal model. The pulsation disappears as the tail-cuff is inflated and restart to pulsate when the tail-cuff is deflated, which equals systolic pressure. The cuff is connected to a tail-cuff sphygmomanometer, and the blood pressure is recorded on a chart (20).
The rats were conscious during measurement, warmed at 28°C for 30 min in a thermostatically controlled heating cabinet (UgoBasille, Italy) to detect tail artery pulse accurately. The tail was passed from the heating cabinet through a miniaturized cuff, and a tail-cuff sensor was connected to an amplifier (ML125 NIBP, AD Instruments, Australia). The amplified pulse was recorded during automatic inflation and deflation of the cuff. Systolic blood pressure was defined as the cuff inflation pressure at which waveform become indistinguishable from baseline noise. The average of at least three measurements was taken at each occasion.
3) Electrocardiogram recordings
Following the systolic blood pressure assessment, the rats were anesthetized again with urethane (0.6 mL/100 gm, b.w., intramuscular), and then the electrocardiogram was recorded.
In vitro tissue processing and studies
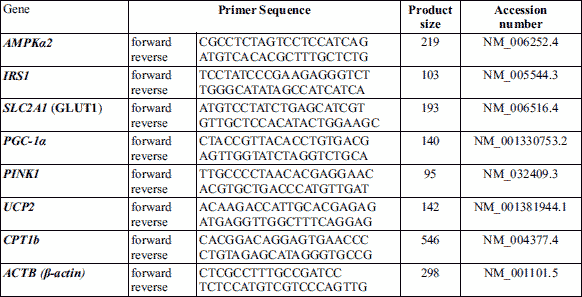
Following the heart’s functional assessment, the rats were sacrificed under ether anesthesia; the thoracic aorta was removed for subsequent evaluation of vascular reactivity. The heart was excised for biochemical measurements (cardiac tyrosine hydroxylase, norepinephrine, glucose transporter 1 (GLUT1), insulin receptor substrate 1 (IRS1), peroxisome proliferator-activated receptor γ co-activator-1 (PPARγ co-activator-1) (PGC-1), phosphatase and tensin homolog-induced kinase 1 (PINK1), carnitine palmitoyltransferase I (CPT1), uncoupling protein 2 (UCP2) and adenosine monophosphate-activated protein kinase alpha 2 (AMPKα2)).
Vascular reactivity studies of isolated aorta
The thoracic aorta was immediately excised and cut into sagittal rings 3 mm in length and transferred to organ chambers rinsed with 10 mL of freshly prepared Krebs-Heinseleit solution (NaCl 118 mM, KCl 4.7 mM, CaCl2 2.5 mM, MgSO4 1.2 mM, NaHCO3 25 mM, KH2PO4 1.2 mM, glucose 10 mM) maintained at 37°C, pH 7.4 and incubated within 95% O2 and 5% CO2. Between every two hooks, the aorta’s rings were mounted, and the hooks attached to an isometric force transducer, which in turn connected to the system of data acquisition (Power lab 8SP, AD Instruments) for consequence recording the tension of 1 gm for 1 h (21). The phenylephrine response curve (10, 20 and 40 µg concentrations) on isolated rat ring was generated from all the groups. Moreover, the response curve of isolated rat’s aortic ring to cumulative doses of acetylcholine can be calculated by the percentage of relaxation induced by 20 µg acetylcholine divide by a submaximal dose of 20 µg phenylephrine-induced contractions for every group (Fig. 1).
Quantitative-RT-PCR
During sacrificing, the hearts were excised and flourished with phosphate buffered-saline and cut into several pieces; part of these cardiac pieces was rinsed in stabilizing reagent RNA later solution, and other pieces were freshly cleaned and then stored at –80°C for subsequent processing. Total RNA extraction was performed from homogenized 25 mg of stored tissues using RNeasy Fibrous Tissue Mini Kit (Qiagen, Hilden, Germany). The RNA concentration was determined using a UV spectrophotometer (Optima SP-3000+, Japan) at 260 and 280 nm. Reverse transcription was performed for 1.5 µg of RNA using Miscript II RTPCR kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. A quantitative RTPCR was performed with qPCR System (Life Technologies, Carlsbad, CA, USA) and QuantiTect SYBR Green Master Mix (Qiagen, Hilden, Germany). All the experiments were performed in triplicate, and the results were normalized to a b-actin housekeeping gene. The sequences of used primers are listed in Table 1.
Fluorometric determination of norepinephrine in the myocardial tissue
The freshly stored cardiac pieces were weighed and homogenized using a Teflon pestle homogenizer (Thomas Scientific, Swedesboro, NJ, USA) in 10 volumes of 5% of trichloroacetic acid. The homogenate was then centrifuged at cooling 15000 rpm in a cooling centrifuge for 15 min. The supernatant was decanted in a chilled clean small tube for epinephrine analysis following the fluorometric method described previously using the Jenway 6200 fluorometer (Jenway Inc., Stone, UK) (22).
Tyrosine hydroxylase (HT) activity
The assay was performed by the simple fluorescence assay technique previously described (23). The dopa enzyme’s rapid isolation was carried out by a double column procedure (the top column of Amberlite CG-50 and Aluminum oxide on the bottom column). The Amberlite CG-50 column was used for erasing the interfering substances, while the second column of aluminum oxide for dopa adsorbance. The dopa was then eluted using 0.5 M acetic acid and determined by the previously described hydroxy indole method (24).
Statistical analysis
The experiments were performed in triplicates. The data were expressed by mean ± standard error (SE) and were statistically analyzed using GraphPad v.6.0.0 software (GraphPad, San Diego, USA). We checked normality of distribution by Shapiro-Wilk test. The data were analyzed using two-way ANOVA and three-way ANOVA (post-hoc Tukey’s test) to compare the mean differences between the experimental groups.
RESULTS
Body wight
At the end of the work, MPTP group showed significant (P < 0.001) wight loss compared to control group. Citicoline therapy to MPTP-treated rats attenuated wight loss however it was not significant effect compared to MPTP group (Table 2).
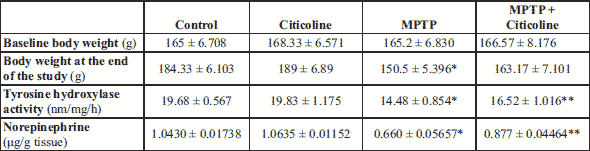
Body weight 1: at the start of the work; body weight 2: at the end of the work.
Modulatory effect of citicoline on biochemical parameters of cardiac tissues
The represented data in Table 2 show that tyrosine hydroxylase and norepinephrine were significantly decreased in the MPTP group compared with the normal control group. The administration of the MPTP-treated group with citicoline at the indicated dose showed a significant (P < 0.001) increased in the indicated markers to the normal group levels compared to the corresponding MPTP control rats.
As represented in Fig. 1, there is a significant (P < 0.001) upregulation in the mRNA levels of UCP2 and AMPKα2 in the MPTP-treated group compared with the normal control group. On the other hand, the inoculation of MPTP-treated rats with citicoline with the indicated dose showed a significant (P < 0.001) downregulation of these markers to the levels near normal rats compared to the corresponding MPTP control rats. Moreover, the mRNA levels of GLUT1, IRS1, PGC1a, and PINK1 showed a significant (P < 0.01) downregulation at the transcriptional level in MPTP-treated rats when compared with the normal control group. On the contrary, there is a significant (P < 0.01) upregulation of mRNA levels of the same markers in the MPTP-treated group with citicoline close to the normal control group’s level compared to the corresponding MPTP control rats. The mRNA relative CPT1 expression showed no significant change among the studied groups. These results are log to the normalized expression values against the beta actin gene.
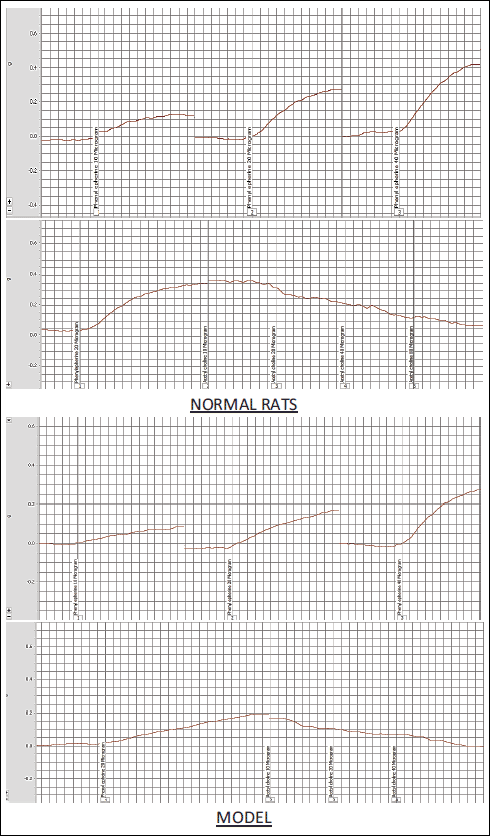 |
Fig. 1. Vascular reactivity studies of isolated aorta. The phenylephrine response curve (10, 20 and 40 µg concentrations) on isolated rat ring was generated from all the groups. Moreover, the response curve of isolated rat’s aortic ring to cumulative doses of acetylcholine on top of submaximal dose of 20 µg phenylephrine-induced contractions for every group. |
Regulatory effect of citicoline on the echocardiography recordings
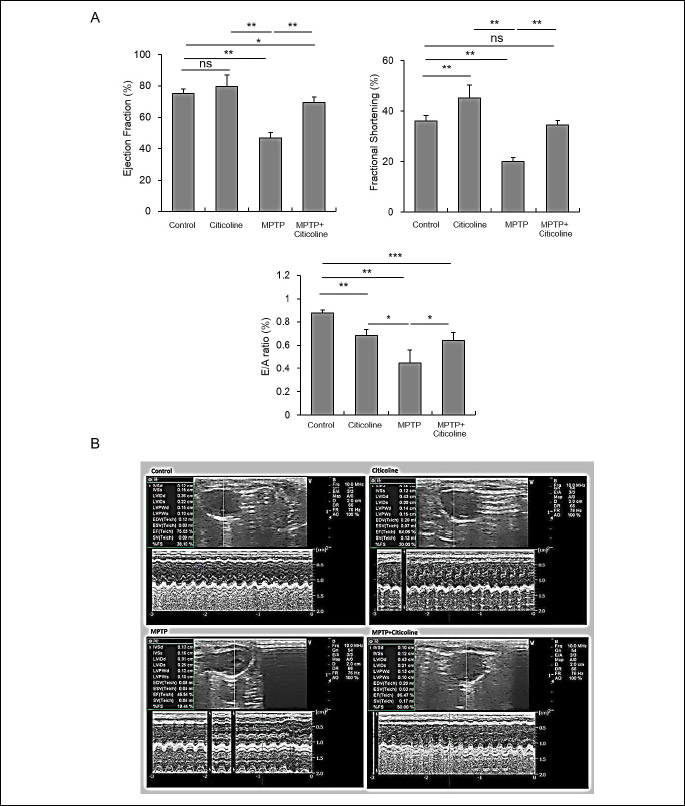
The data presented in Table 3 and Fig. 3 show the relevant echocardiography readings during the heart’s functional assessment. There was a significant (*P < 0.01) reduction of EF, FS, E/A ratio, LVDd, LVPWD ,HR, SV and CO in MPTP-treated rats when compared to normal control. Administration of citicoline significantly (**P < 0.01) increased EF, FS, LVDd, HR, SV and CO compared to MPTP group. No significant difference was found in LVSd and LVPWS between the studied groups.
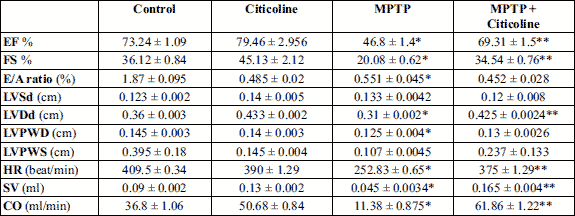
Abbreviations: EF, ejection fraction; FS, fractional shortening; LVDd, left ventricular internal diameter during diastole; LVSd, left ventricular internal diameter during systole; LVPWD, left ventricular posterior wall thickness during diastole; LVPWS, left ventricular posterior wall thickness during systole; IVSS, interventricular septal wall thickness during systole; IVSD, interventricular septal wall thickness during diastole; CO, cardiac output.
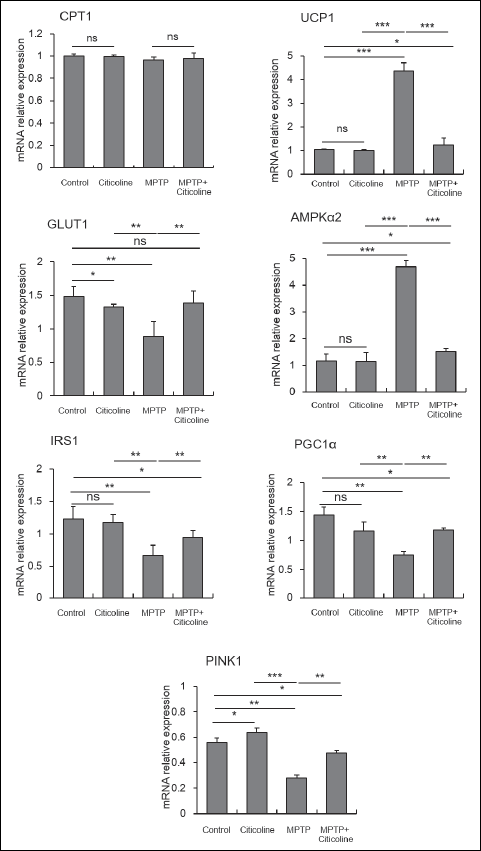 |
Fig. 2. Modulatory effect of citicoline on transcriptional levels of MPTP-treated cardiac tissue rats. Cardiac tissues of MPTP-exposed rats showed significant (***P < 0.001) upregulation of UCP2 and AMPKα2, then significant (**P < 0.01) downregulation of GLUT1, IRS1, PGC1α, and PINK1 were also observed when compared to the normal control group. On the other hand, citicoline administration to MPTP-treated rats was significantly (**P < 0.01) modulated to be close to normal compared to MPTP control rats. In contrast, the mRNA of CPT1 expression was not different between the studied groups. The data were presented as mean ± SE. |
Regulatory effect of citicoline on the electrocardiography recordings
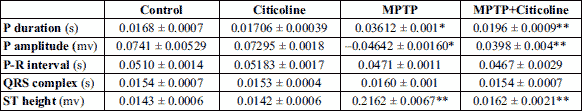
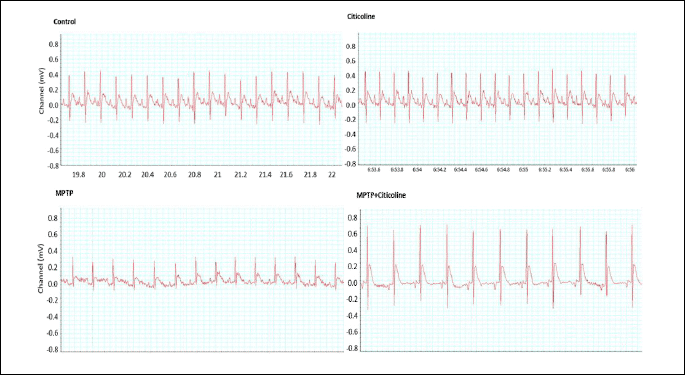
The data presented in Table 4 and Fig. 4 showed a significant (P < 0.01) prolongation of P-wave duration and a decrease of P amplitude in the MPTP treated group compared to the corresponding normal control group. The inoculation of the MPTP-treated group with the indicated dose of citicoline significantly (P < 0.001) recovered the previous events by shortening the P-wave duration and increasing the P amplitude close to the normal group when compared to the MPTP-treated group. The P-R interval and QRS complex showed no significant changes among the studied groups. Despite the significant (P < 0.001) elevation in the height of the ST segment from the isoelectric baseline in MPTP-treated rats, there is a significant (P < 0.001) amelioration beyond the administration of citicoline to the MPTP-treated rats with the indicated dose to be close to the normal control group.
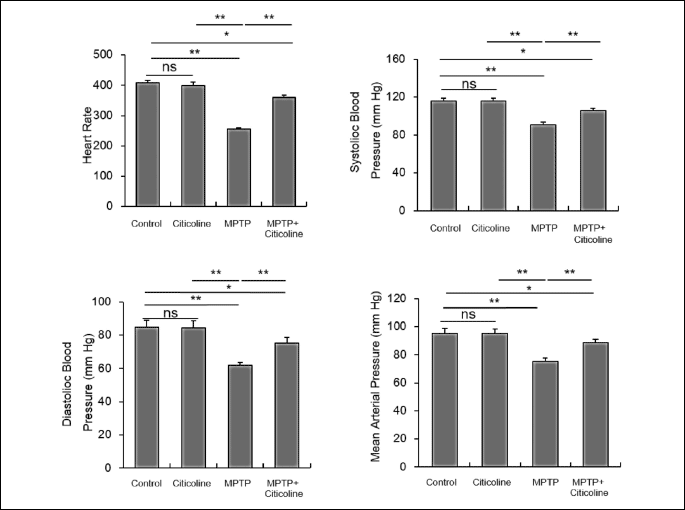
Ameliorative effect of citicoline on the arterial blood pressure
The presented data in Figure 5 showed a significant (P < 0.001) depletion of systolic, diastolic blood pressure, heart rate, and mean arterial blood pressure in the MPTP-exposed group when compared with the normal control group. Interestingly, citicoline administration to the MPTP-exposed rats revealed a significant (P < 0.001) increase of the parameters compared to MPTP and for the mean and diastolic blood pressures the values in citicoline + MPTP group were similar to the control group.
Impact of citicoline on vascular reactivity-based aortic rings contractions
The vascular reactivity of the aortic rings’ contraction in response to various phenylephrine doses has been presented in Fig. 5. The results indicated a significant (P < 0.001) decrease in the contraction force in the MPTP-treated group at doses 10, 20, and 40 µg of phenylephrine when compared to the normal control group. On the other hand, there is a significant (P < 0.001) recovery of the contraction force of the aortic ring contraction at the indicated doses of phenylephrine in the MPTP-treated group with citicoline near to normal levels of the control group. There is a significant elevation in the aortic rings’ contraction force in a dose-dependent manner in all the studied groups.
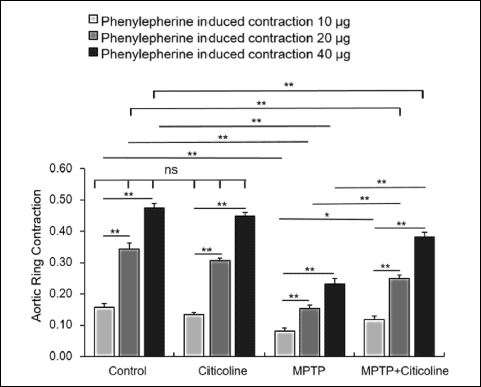 |
Fig. 6. Regulatory effect of citicoline on aortic ring contraction of MPTP-treated rats. The contraction force of aortic rings showed a significant (**P < 0.01) decrease in MPTP-exposed rats at the indicated doses of phenylephrine compared to the normal control group. On the contrary, the citicoline administration of MPTP-exposed rats showed a significant (**P < 0.01) recovery close to normal control rats. The data were presented as mean ± SE. |
DISCUSSION
The present study investigated the cardiovascular system’s chemical and mechanical dysfunctions during the experimental model of dysautonomia-induced rats using MPTP. Previous studies used an experimental model of Parkinson’s disease by using MPTP (25, 26). The main chemical dysfunction observed in the cardiovascular myocytes of MPTP-treated rats were depletion of tyrosine hydroxylase activity, norepinephrine secretion, decrease the relative mRNA transcription of GLUT1, IRS1, PPARγ co-activator-1, PGC-1, PINK1. Furthermore, and increased the relative mRNA transcription of uncoupling protein 2 (UCP2) and adenosine monophosphate-activated protein kinase alpha 2 (AMPKα2). Moreover, the significant mechanical dysfunctions recorded in cardiac myocytes of MPTP-treated rats were reduced fractional cell shortening, prolongation of P-wave duration relaxation, and decreased velocities of both shortening and re-lengthening. The administration of citicoline has modulated the above abnormalities.
The evidence of the anatomical studies suggested that the cardiovascular system’s sympathetic innervation is affected in neurodegenerative disorders, including synucleinopathies, which develop Lewy body’s formation, and Parkinson’s disease (27). The current study documented that the cardiac system’s sympathetic denervation occurred in MPTP-treated rats (28) and was modulated upon administration of citicoline. Other previous studies confirmed our results that shown depletion of cardiac norepinephrine in MPTP-treated rats (29). Ren et al., studies (25) demonstrated that the number of β-adrenergic receptors is likely to share the decreased adrenergic responsiveness and eventually impair mechanical cardiac functions in MPTP-treated mice. Even though sympathetic denervation may lead to postsynapses’ supersensitivity due to lack of neural uptake of norepinephrine, various denervation-based-supersensitivity models remain controversial. Besides, Amino et al., (30) demonstrated that the cardiac nerve fibers and terminals were morphologically well preserved in the MPTP-treated mice, despite cardiac sympathetic dysfunction indicated by the significant decreases in the contents of cardiac noradrenaline and dopamine. Furthermore, MPTP fundamental impact focuses on its ability to cross the blood-brain barrier after activation to MPP+, and this active form selectively enters the postganglionic sympathetic nerves through monoamine transporter (9). The MPP+ eventually recruited in mitochondria resulted in catecholamine neurodegeneration.
It is noteworthy that citicoline administration regulated cardiac catecholamine levels near the normal control, matched with previous investigations (31). The tyrosine hydroxylase activity in the dopaminergic neurons has been reduced in MPTP-treated rats and then increased near to normal level upon citicoline administration. The critical effect of citicoline could be due to the regulation of neuroinflammatory signaling.
Multiple studies reported that citicoline protects from heart damage caused by ischemia/reperfusion processes by reducing mitochondrial permeability. The results indicated the regulation of oxidative phosphorylation and cis-aconitase mitochondrial enzyme, which results in avoidance of oxidative disruption of mitochondrial DNA and the release of cytokines (32). The protective effect of citicoline upon the dopaminergic system has been investigated previously based on its pharmacological actions (33).
The most dominant heart problems found in the current research were decreased cardiac contraction and P waves’ prolongation during the time of relaxation, associated with the reduced maximal velocity of contraction/relaxation in MPTP-treated rats. Moreover, depletion of systolic, diastolic blood pressure, heart rate, and mean arterial blood pressure in MPTP-exposed rats were also observed. These results were confirmed by previous studies that reported that changes of heart rate variability through electrocardiographic assessment in Parkinson’s disease model was used MPTP (34, 35). Various factors may have disrupted the heart’s mechanical functions, including impaired functions of contractile proteins like actin and myosin isoforms, decreased availability of intracellular Ca2+ due to affected myofilament Ca2+ sensitivity, despite other alternatives that may exist but remain to be unclear.
The electrocardiogram can also be used in isolated Langendorff rat hearts, as demonstrated by Regev et al. (36) in their study that evaluated the ventricular arrhythmia and the interobserver agreement on ventricular fibrillation. Furthermore, Bernikova et al. (37) evaluated the electrical activity by EEG in reperfusion arrhythmogenesis and correlated it to the duration of repolarization using tetraethylammonium (TEA) that blocks outward potassium currents and pinacidil that opens ATP-sensitive potassium channels.
The current study showed evidence of cardiac function dysregulation using the MPTP-animal model. Upregulation of mitochondrial protein; uncoupling protein 2 family observed in MPTP-treated rats due to excessive production of reactive oxygen species which activated UCP2 transcription to regulate and control the excessive ROS production through negative feedback (38). The function of AMPK in the heart is not cleared yet and become active during stress-induced hypertrophy. The present investigation showed upregulation of the AMPKα2 transcription in the MPTP-treated group. Furthermore, the increasing of AMPKα2 mediates the adiponectin’s antihypertrophic effects (39). The AMPK can also influence the metabolic activity of gene expression. It plays a role in skeletal muscle exercise by increasing the peroxisome proliferator activated-receptor γ-coactivator-1α (PGC-1α) similar to what happened in our study; it was increased to upregulate the transcription of PGC-1α and Glut1 to regulate the mitochondrial oxidation pathways (40).
In the same perspective of observing metabolic and inflammatory markers in cardiovascular diseases Badacz et al. (41) showed in their study an association between proinflammatory cytokines, chemokines and the atherosclerotic plaque with internal carotid artery stenosis.
Citicoline is involved in patients’ protection of memory disorders against the excessive generation of oxidative stresses. In the present study, citicoline contributed to cardiac recovery when administered to MPTP-treated rats. The citicoline functions improved the mitochondrial dysfunctions and mitophagy by ameliorating UCP2, AMPK-α2 transcriptions (42). Several studies introduced valuable evidence on citicoline’s protective effect upon myocardial dysfunction during the rats’ heart reperfusion (25). Similarly, citicoline proved a protective function against calcium retention through modulation of PGC-1α, which regulates the oxidative metabolism and biogenesis of mitochondria for cardiac dysautonomia (43).
Additional studies demonstrated that citicoline aid in the synthesis of phosphatidylcholine that is a vital structure of cell membranes and mediators of cell signaling. Another likely mechanism of the protective effect of citicoline in the present study is its action to block inflammation by inhibiting phospholipase A2. This enzyme is involved in the breakdown of membrane phospholipids into arachidonic acid. The oxidative metabolism of arachidonic acid contributes to the generation of neuroinflammation and reactive oxygen species (ROS). By blocking phospholipase A2, citicoline may contribute to the reduction of inflammation, ROS formation, and neuronal damage (44). Citicoline causes an increase in SIRT1 (silent information regulator 1, SIRT1). SIRT1 belongs to the histone deacetylase family and regulates metabolic homeostasis and neuronal aging (45). In the cardiovascular system, activation of SIRT1 can not only protect at the cellular level against oxidative stress, but also offer increased survival at the systemic level to limit coronary heart disease and cerebrovascular disease (46).
The present study evaluated the effect of citicoline therapy on cardiovascular function and investigated some biochemical markers’ levels. Citicoline improved cardiac contractility, electrical activity, blood pressure, and vascular reactivity. Citicoline also increased cardiac norepinephrine and tyrosine hydroxylase and improved markers related to ROS scavenger, mitochondrial permeability, calcium homeostasis on the cellular level, metabolic homeostasis, and mitochondrial biogenesis. Further studies are needed using electron microscopy and investigating other levels as post transcriptional or translational levels required to validate the changes of the metabolic markers (UCP2, AMPKα2, GLUT1, IRS1, PGC1α, PINK1, and CPT1) through immunohistochemistry or Western blot analysis for better evaluation of the citicoline’s cellular and molecular effects on cardiovascular autonomic dysfunction.
Availability of data: The data that support the findings of this study are available from the corresponding author (S.N. Amin).
Acknowledgement: We would like to thank Dr. Ahmed Omar Elkady (M.Sc. cardiology, Military medical academy, Egypt) for his kind help in echocardiography.
Source of funding: No fund received for the present work.
Conflict of interests: None declared.
REFERENCES
- Loewy AD, Spyer LK. Central Regulation of Autonomic Functions. New York, Oxford University Press, 1990.
- Higgins CB, Vatner SF, Braunwald E. Parasympathetic control of the heart. Pharmacol Rev 1973; 25: 119-155.
- Jain S, Goldstein DS. Cardiovascular dysautonomia in Parkinsons disease: from pathophysiology to pathogenesis. Neurobiol Dis 2012; 46: 572-580.
- Goldstein DS. Dysautonomia in Parkinson’s disease: neurocardiological abnormalities. Lancet Neurol 2003; 2: 669-676.
- Post KK, Singer C, Papapetropoulos S. Cardiac denervation and dysautonomia in Parkinson’s disease: a review of screening techniques. Parkinsonism Relat Disord 2008; 14: 524-531.
- Sundstrom E, Stromberg I, Tsutsumi T, Olson L, Jonsson G. Studies on the effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) on central catecholamine neurons in C57BL/6 mice. Comparison with three other strains of mice. Brain Res 1987; 405: 26-38.
- Przedborski S, Jackson-Lewis V, et al. Unilateral MPTP-induced parkinsonism in monkeys. A quantitative autoradiographic study of dopamine D1 and D2 receptors and re-uptake sites. Neurochirurgie 1991; 37: 377-382.
- Liu X, Wei B, Bi Q, et al. MPTP-induced impairment of cardiovascular function. Neurotox Res 2020; 38: 27-37.
- Fukumitsu N, Suzuki M, Fukuda T, Kiyono Y, Kajiyama S, Saji H. Reduced 125I-meta-iodobenzylguanidine uptake and norepinephrine transporter density in the hearts of mice with MPTP-induced parkinsonism. Nucl Med Biol 2006; 33: 37-42.
- Natale G, Kastsiushenka O, Fulceri F, Ruggieri S, Paparelli A, Fornai F. MPTP-induced parkinsonism extends to a subclass of TH-positive neurons in the gut. Brain Res 2010; 1355: 195-206.
- Algeri S, Ambrosio S, Garofalo P, Gerli P. Peripheral effects of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and its main metabolite 1-methyl-4-phenylpyridinium ion (MPP+) in the rat. Eur J Pharmacol 1987; 141: 309-312.
- Sbardella D, Coletta A, Tundo GR, et al. Structural and functional evidence for citicoline binding and modulation of 20S proteasome activity: novel insights into its pro-proteostatic effect. Biochem Pharmacol 2020; 177: 113977. doi: 10.1016/j.bcp.2020.113977
- Grieb P. Beneficial effects of exogenous CDP-choline (citicoline) in EAE. Brain 2015; 138: e388. doi: 10.1093/brain/awv140
- Secades J, J, Dlvarez-Sabin J, Rubio F, Lozano R, Davalos A, Castillo J. Citicoline in intracerebral hemorrhage: a double-blind, randomized, placebo-controlled, multi-centre pilot study. Cerebrovasc Dis 2006; 21: 380-385.
- Turkkan A, Alkan T, Goren B, Kocaeli H, Akar E, Korfali E. Citicoline and postconditioning provides neuroprotection in a rat model of ischemic spinal cord injury. Acta Neurochir (Wien) 2010; 152: 1033-1042.
- Salamah A, Mehrez M, Faheem A, El Amrousy D. Efficacy of citicoline as a neuroprotector in children with post cardiac arrest: a randomized controlled clinical trial. Eur J Pediatr 2021; 180: 1249-1255.
- Safavi M, Hosseini-Sharifabad A, Seyed-Yousefi Y, Rabbani M. Protective effects of citicoline and benfotiamine each alone and in combination on streptozotocin-induced memory impairment in mice. Clin Psychopharmacol Neurosci 2020; 18: 81-92.
- Zhang Q, Heng Y, Mou Z, et al. Reassessment of subacute MPTP-treated mice as animal model of Parkinson’s disease. Acta Pharmacol Sin 2017; 38: 1317-1328.
- Watson CJ, Hunt F, Messer S, et al. In situ normothermic perfusion of livers in controlled circulatory death donation may prevent ischemic cholangiopathy and improve graft survival. Am J Transplant 2019; 19: 1745-1758.
- Wilde E, Aubdool AA, Thakore P, et al. Tail-cuff technique and its influence on central blood pressure in the mouse. J Am Heart Assoc 2017; 6: e005204. doi: 10.1161/JAHA.116.005204
- Csanyi G, Lepran I, Flesch T, Telegdy G, Szabo G, Mezei Z. Lack of endothelium-derived hyperpolarizing factor (EDHF) up-regulation in endothelial dysfunction in aorta in diabetic rats. Pharmacol Rep 2007; 59: 447-455.
- White CE. Fluorometric analysis. Anal Chem 1960; 32: 47-53.
- Nagatsu T, Oka K, Numata (Sudo) Y, Kato T. A simple and sensitive fluorescence assay for tyrosine hydroxylase activity. Anal Biochem 1979; 93: 82-87.
- Johansson B, Roos. B-E. 5-Hydroxyindole acetic acid and homovanillic acid in cerebrospinal fluid of patients with neurological diseases. Eur Neurol 1974; 11: 37-45.
- Ren J, Porter JE, Wold LE, Aberle NS, Muralikrishnan D, Haselton JR. Depressed contractile function and adrenergic responsiveness of cardiac myocytes in an experimental model of Parkinson disease, the MPTP-treated mouse. Neurobiol Aging 2004; 25: 131-138.
- Rodriguez-Cruz A, Romo-Mancillas A, Mendiola-Precoma J, Escobar-Cabrera JE, Garcia-Alcocer G, Berumen LC. Effect of valerenic acid on neuroinflammation in a MPTP-induced mouse model of Parkinson’s disease. IBRO Rep 2019; 8: 28-35.
- Iwanaga K, Wakabayashi K, Yoshimoto M, et al. Lewy body-type degeneration in cardiac plexus in Parkinson’s and incidental Lewy body diseases. Neurology 1999; 52: 1269-1271.
- Takatsu H, Nishida H, Matsuo H, et al. Cardiac sympathetic denervation from the early stage of Parkinson’s disease: clinical and experimental studies with radiolabeled MIBG. J Nucl Med 2000; 41: 71-77.
- Amino T, Orimo S, Itoh Y, Takahashi A, Uchihara T, Mizusawa H. Profound cardiac sympathetic denervation occurs in Parkinson disease. Brain Pathol 2005; 15: 29-34.
- Amino T, Uchihara T, Tsunekawa H, et al. Myocardial nerve fibers are preserved in MPTP-treated mice, despite cardiac sympathetic dysfunction. Neurosci Res 2008; 60: 314-318.
- Przedborski S, Kostic V, Jackson-Lewis V, et al. Transgenic mice with increased Cu/Zn-superoxide dismutase activity are resistant toN-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dineinducedneurotoxicity. J Neurosci 1992; 12: 1658-1667.
- Hernandez-Esquivel L, Pavon N, Buelna-Chontal M, Gonzalez-Pacheco H, Belmont J, Chavez E. Citicoline (CDP-choline) protects myocardium from ischemia/reperfusion injury via inhibiting mitochondrial permeability transition. Life Sci 2014; 96: 53-58.
- Mao J, Liu S, Fu C. Citicoline retards myopia progression following form deprivation in guinea pigs. Exp Biol Med (Maywood) 2016; 241: 1258-1263.
- Rahman F, Pechnik S, Gross D, Sewell L, Goldstein DS. Low frequency power of heart ratevariability reflects baroreflex function, not cardiac sympathetic innervation. Clin Auton Res 2011; 21: 133-141.
- Valappil RA, Black JE, Broderick MJ, et al. Exploring the electrocardiogram as a potential tool to screen for premotor Parkinson’s disease. Mov Disord 2010; 25: 2296-2303.
- Regev A, Takacs H, Farkas AS, et al. Application of ventricular tachyarrhythmia definitions of the updated Lambeth Conventions provides incompatibility with earlier results, masks antifibrillatory activity and reduces inter-observer agreement. J Physiol Pharmacol 2019; 70: 37-49.
- Bernikova OG, Sedova KA, Durkina AV, Azarov JE. Managing of ventricular reperfusion tachyarrhythmias - focus on a perfused myocardium. J Physiol Pharmacol 2019; 70: 757-763.
- Azzu V, Brand MD. The on-off switches of the mitochondrial uncoupling proteins. Trends Biochem Sci 2010; 35: 298-307.
- Shibata R, Ouchi N, Ito M, et al. Adiponectin-mediated modulation of hypertrophic signals in the heart. Nat Med 2004; 10: 1384-1389.
- Reznick RM, Shulman GI. The role of AMP-activated protein kinase in mitochondrial biogenesis. J Physiol 2006; 574: 33-39.
- Badacz R, Podolec J, Przewlocki T, et al. The role of chemokine CCL5/RANTES and metalloproteinase-9 as inflammatory modulators in symptomatic internal carotid artery stenosis. J Physiol Pharmacol 2019; 70: 545-555.
- Synoradzki K, Grieb P. Citicoline: a superior form of choline? Nutrients 2019; 11: 1569. doi: 10.3390/nu11071569
- Yilmaz M, Coskun C, Yalcin M, Savci V. CDP-choline prevents cardiac arrhythmias and lethality induced by short-term myocardial ischemia-reperfusion injury in the rat: involvement of central muscarinic cholinergic mechanisms. Naunyn-Schmiedeberg’s Arch Pharmacol 2008; 378: 293-301.
- Adibhatla RM, Hatcher JF. Citicoline decreases phospholipase A2 stimulation and hydroxyl radical generation in transient cerebral ischemia. J Neurosci Res 2003; 73: 308-315.
- Hurtado O, Hernandez-Jimenez M, Zarruk JG, et al. Citicoline (CDP-choline) increases Sirtuin1 expression concomitant to neuroprotection in experimental stroke. J Neurochem 2013; 126: 819-826.
- Chong ZZ, Wang S, Shang YC, Maiese K. Targeting cardiovascular disease with novel SIRT1 pathways. Future Cardiol 2012; 8: 89-100.
A c c e p t e d : February 26, 2021
ORCID: 0000-0001-9232-2389