Ototoxicity, i.e., cochlear and/or vestibular toxicity, was first seen in individuals who used streptomycin and dihydrostreptomycin for the treatment of tuberculosis (6). After 1963 when gentamicin first became available, it has remained the most widely-used aminoglycoside (7, 8). While gentamicin is a highly effective treatment for idiopathic endolymphatic hydrops (Menieres disease), a third of the patients who use the drug develop hearing loss (9). The bactericidal actions of the aminoglycosides involve a series of defined steps including their binding to ribosomes and the inhibition of protein synthesis (10).
Cisplatin, a widely-used chemotherapeutic agent but not an aminoglycoside, also has toxic actions, which limits the dose at which it is given. The negative side effects of this drug are similar to those of the aminoglycosides at the level of the inner ear.
Tinnitus, also commonly referred to as ringing in the ears, is distracting and occasionally debilitating to the extent it may cause sleep loss. Many agents are believed responsible for causing tinnitus, e.g., excessively loud noise, medications, etc., and the currently-available treatments are less than optimally effective in reducing tinnitus.
Each of these subjects will be discussed in the current survey. The amount of information related to the protective effects of melatonin in these conditions varies widely with the bulk of data describing the benefits of the indoleamine against aminoglycoside toxicity.
Aminoglycoside molecules per se are not inherently toxic; rather, they require the redox capacity of a transition metal ion to become destructive (1). Thus, an aminoglycoside chelates a metal ion to generate chelated metal complexes (11, 12) which are redox active and, therefore, give rise to reactive oxygen (ROS) including free radicals. Once produced, these oxidizing agents have the capability of mutilating nearby biomolecules (13). There is direct evidence that at least some of the damage inflicted by aminoglycosides involves ROS and reactive nitrogen species (RNS) (14-16). The generated toxic agents not only cause molecular damage but also initiate cell death via apoptosis and necrosis.
There is experimental evidence supporting the role of free radicals and related reactants in ototoxicity that results from the interaction of an aminoglycoside with cellular molecules. Hence, animals over expressing cytosolic superoxide dismutase (SOD), the enzyme that metabolizes the superoxide anion radical (O2
Since aminoglycosides react with transition metals, e.g., iron, to generate destructive ROS/RNS, it was presumed that metal chelators may be effective in ameliorating loss of hair cells caused by these drugs. Indeed, the use of iron chelators, i.e., desferoxamine or dihydroxybenzoate, do reduce ototoxicity that results from aminoglycoside treatment; this is accomplished without interfering with the antibiotic activity of the drug (2).
There is also a modicum of evidence indicating that aspirin (sodium salicylate) attenuates gentamicin-mediated ototoxicity (5). In a double-blind study in which gentamicin treatment was coupled with aspirin administration, a reduced incidence of hearing loss was observed. Aspirin usage, however, was associated with gastric bleeding in a small percentage of the patients. Mechanistically, it was presumed that either the metal chelating or antioxidant activities of aspirin accounted for its protective actions against gentamicin in this clinical trial. The use of other antioxidants in an attempt to alleviate the loss of inner ear hair cells during aminoglycoside administration has been attempted with variable success. The antioxidants include d-methionine and lipoic acid.
Melatonin, N-acetyl-5-methoxytryptamine, is an endogenously produced indoleamine that is best known for its ability to synchronize circadian and circannual rhythms (21, 22). While it was initially discovered as a secretory product of the pineal gland of vertebrates (23), it is in fact produced in all organisms of the animal kingdom from unicells to humans (24) and also in plants (25-27). Also, although discovered as a synthetic product of tryptophan metabolism in the mammalian pineal gland, it is now known to be produced in a variety of different cells/tissues (28), including in the cochlea (29). During evolution, melatonins original function seems to have been as an antioxidant with its other functions being later additions to its physiological repertoire (28).
 |
Fig. 1. Proposed mechanism of aminoglycoside (AG) toxicity at the level of the hair cell in the cochlea and utricle of the inner ear. After the AG enters the hair cell through mechano-electrical transducer channels, it forms a complex with a transition metal, in this case iron (Fe). These redox active complexes generate ROS/RNS including the superoxide anion radical, hydrogen peroxide, hydroxyl radical, peroxynitrite anion, etc. These reactants activate JNK (c-Jun N-terminal kinase) which then translocate (straus locates) to the nucleus to activate genes in the cell death pathway; these products are then transferred to the mitochondria (mito) where they promote the release of cytochrome c (Cyt c). In the cytosol, Cyt c triggers the activation of a series of caspases followed by apoptosis (programmed cell death) via what is referred to as caspase-dependent cell death. In addition to this caspase-dependent apoptotic pathway, AG may also kill cells via caspase-independent mechanisms. |
Melatonin also is an uncommonly potent direct free radical scavenger and indirect antioxidant (30, 31). Moreover, in the process of scavenging toxic free radicals, melatonin is converted to metabolites that seem to be as least as effective as melatonin itself in neutralizing free radicals and associated toxic agents. These melatonin metabolites include cyclic 3-hydroxymelatonin (32), N1-acetyl-N2-formyl-5-methoxykynuramine (AFMK) (33), N1-acetyl-5-methoxykynuramine (AMK) (34) and perhaps others (35). The functional diversity of melatonin and its metabolites is emphasized by its ability to neutralize toxic derivatives of oxygen which include the most highly reactive products, the hydroxyl radical (
In addition to its ability to directly detoxify radicals, melatonin has a variety of other means by which it quells oxidative damage. Notably, melatonin stimulates antioxidative enzymes which convert free radicals to innocuous non-reactive products (31, 40, 41), it may chelate transition metals which normally would participate in redox reactions (42), it reduces election leakage from mitochondria thereby preventing free radical formation (38, 43-45) and it promotes the synthesis of another important antioxidant, glutathione (46). Finally, melatonin and/or AMK inhibit the prooxidative enzyme, nitric oxide synthase (47). This enzyme converts arginine to nitric oxide making it available to couple with the O2
The combination of actions of melatonin, as described above, in resisting oxidative/nitrosative stress presumably make the indoleamine an important antioxidant. In comparative studies with other well-known free radical scavenges, e.g., vitamins C and E, etc., melatonin has often been found to be better in terms of its ability to protect against molecular destruction by toxic oxygen derivatives (48). One of melatonins advantages is seemingly its ability to enter both the lipid (for example, membranes) and aqueous (for example, cytosol and nucleoplasm) environments of the cell where it scavenges radicals and their derivatives. Often the unique solubility of antioxidants (for example, vitamin E works exclusively in the lipid portions of cells) limits their efficacy in neutralizing radicals that are produced in all subcellular compartments. Additionally, the actions of melatonin at the level of the mitochondria (43, 45, 49-51), which are normally the major site of intracellular free radical generation, may well contribute to its high efficacy in protecting against free radical damage.
In the organ of Corti of the cochlea, the outer cells are crucial elements of the transduction mechanism by which environmental sound is converted to an electrical signal that is sent to the brain via the cochlear division of cranial nerve VIII. The hair cells, when transducing environmental sound waves, produce what is referred to as otoacoustic emissions (OAEs) (52). The OAEs are recorded by placing an external probe in the auditory canal of the temporal bone. This technique allows the detection of electrical signals representing sound using an easy and non-invasive method (53). OAEs are subclassified according to the type of acoustic stimulation used to produce them; one subcategory of OAEs is distortion products, or DPOAEs (54).
Realizing that the antioxidant melatonin is present in the cochlea (55) and that excessive free radical generation compromises the function of cochlear outer hair cells and, therefore, the recordable OAEs, Lopez-Gonzalez and colleagues (56) tested whether melatonin administration would alter the DPOAEs from hair cells experiencing excessive oxidative stress due to ischemia. In this study, melatonin (250 µg) or a mixture of other antioxidants consisting of 0.25 mg
When the organ of Corti is rendered ischemic as a result to the death of the animal, the cells generate free radicals due to hypoxia and DPOAEs can be recorded. The fact that both melatonin and the antioxidant mixture prolonged the duration of the DPOAEs indicates that they successfully scavenged the radicals produced and slowed the death of the hair cells. Lopez-Gonzalez et al. (56) surmised, based on the doses of melatonin (250 µg) and antioxidant mixture (64.25 mg) administered, that melatonin was roughly 350 times more effective than the mixture in terms of their abilities to reduce free radical formation. Since both antioxidant concoctions seem to have been maximally effective in limiting free radical generation, however, more studies are required to determine the relative efficacies of these antioxidants.
In a related study, Lopez-Gonzalez and co-workers (16) also examined whether melatonin would influence the recorded DPOAEs measured after rats were treated with 5 daily intramuscular injections of either gentamicin (160 mg/kg body weight (BW)) or tobramycin (200 mg/kg BW). Melatonin was either given in the drinking water (10 mg/L) or as a daily subcutaneous (250 µg) or intramuscular (250 µg) injection. The ototoxicity measured as DPOAEs which accompany the administration of the aminoglycosides was maximal at 3-5 days during treatment; the DPOEAs disappeared 9 days after aminoglycoside administration was discontinued. By contrast, normal OEAs reappeared within 5 days when either gentamicin or tobramycin were given in combination with melatonin. Also, importantly, melatonin did not interfere with the antibiotic activities of the aminoglycosides as measured using antibiograms after the inoculation of either Escherichia coli or Pseudomonas aeruginosa.
Since it appears that excessive free radical generation is, at least in part, responsible for the ototoxicity of gentamicin and tobramycin, it is reasonable to assume that the ability of melatonin to function as an antioxidant may well account for its efficacy to reduce DPOEAs generated at the level of the cochlear hair cells. Considering the multiple means by which melatonin functions (as summarized above) to overcome free radical production and the associated molecular damage, which of melatonins effects explain its protective actions in hair cells remains unknown.
At least as devastating as cochlear hair cell loss resulting from the use of aminoglycoside antibiotics, is the irreversible damage to the vestibular hair cells. The vestibular dysfunction that arises is sometimes unrecognized as being related to aminoglycoside use due to the sudden onset of this serious adverse side effect. Moreover, the emotional distress and financial impact related to an episode of vestibular malfunction due to aminoglycoside use may be substantial. Hence, damage to the vestibular apparatus may prolong hospitalization as well as physiotherapy and rehabilitation, but has even more long-range consequences in terms of difficulty in securing or maintaining employment and inability to operate a vehicle (57).
The hair cells of the vestibular apparatus of the inner ear are a major target for the damage inflicted by aminoglycosides. There is general agreement in the published literature that vestibulotoxicity, like ototoxicity, related to aminoglycosides stems, at least in part, from the excessive generation of toxic free radicals in the hair cells (58); radicals not only damage these cells but often cause them to undergo apoptosis (59). Once lost, these cells cannot be replaced so the damage and associated vestibular dysfunction are permanent.
The initiation of free radical generation presumably involves the natural action of aminoglycosides to chelate iron which greatly enhances the likelihood of the hair cells to produce toxic oxygen-based radicals in excess (14, 56, 60, 61). Considering the processes described, Song and colleagues (62) found that the administration of an iron chelator to animals receiving aminoglycosides limited vestibular toxicity without changing the antibacterial properties of the drug.
Within the vestibular apparatus, the damage caused by the aminoglycosides is not uniformly distributed. Rather, damage initially develops at the top of the cristae and at the striolar regions of the maculi; it then progresses to the type I hair cells and later to the type II cells (63).
While all currently-used aminoglycosides have been found to cause vestibulotoxicity by destroying hair cells in the ampullae cristae (58, 61-63), there does seem to be some organ specificity with gentamicin seemingly being more destructive to the hair cells of the vestibule than those of the cochlea while the actions of amikacin are opposite to this (64). It was initially thought that the concentration of the specific drug within the fluid of the vestibular apparatus was a contributor to the resulting toxicity. Research has not borne this out, however, with the degree of damage to the hair cells being independent of the concentration of the aminoglycosides in the vestibular fluid (14, 65).
Since vestibular hair cell loss may involve the generation of ROS and apoptosis after gentamicin administration, Kim and colleagues (66) reasoned that the antioxidant melatonin may be useful in combating the destruction of the hair cells of the utricles of the rat. For this ex vivo study, utricles were obtained from the temporal bones of 2-4 day old Sprague-Dawley rats; thereafter, the explants were cultured in Dulbeccos Modified Eagles Medium containing 1 mM gentamicin and melatonin at concentrations of either 10, 50 or 100 µM. Following the various treatments, the number of phalloidin-fluorescein isothiocyanate (phalloidin-FITC)-stained hair cells were counted in the utricles. In a dose-response manner, melatonin reduced live hair cell loss although even at the dose of 200 µm, it did not totally reverse the toxicity of 1 mM gentamicin. With the aid of the fluorescent indicator, hydrofluorescent diacetate acetyl ester, the generation of ROS was estimated and found to be significantly elevated in the utricles that were treated with gentamicin only. When the drug was combined with 50 µM melatonin, the fluorescent signal identifying ROS was weakened indicating fewer radicals being formed. Likewise, caspase-3 (estimated using CaspGLOLO fluorescein) activation was elevated in the utricles treated with gentamicin alone and reduced when the aminoglycoside was given in combination with 50 µM melatonin. Western blot analysis confirmed caspase-3 activation by gentamicin and its reduction following treatment with melatonin. In this study, Kim and co-workers (66) did not test the efficacy of the highest dose (100 µm) of melatonin in altering ROS production or caspase-3 activation.
The results of this report, which used explants of newborn rat utricles, documented the ability of melatonin to attenuate loss of hair cells, ROS generation and activation of caspase-3. The implication of these findings is that melatonin may have utility in preventing utricle hair cell loss that results from aminoglycoside toxicity in vivo. Kim et al. (66) concluded that melatonin does prevent oxidative damage and apoptosis in utricular hair cells. Since the toxicity of aminoglycosides may rely on their ability to generate excessive ROS, it is assumed that melatonin could be effective against each of these drugs in reducing utricular hair cell loss.
There is ample justification for the assumption that melatonin may also be capable of overcoming hearing loss due to cisplatin treatment. Cisplatin, a common cancer therapeutic drug, is not an aminoglycoside but it has some similar side effects. Also, like the aminoglycoside, cisplatin is often dose-limited by its toxicity which includes damage to the inner ear (67, 68); one of the underlying mechanisms of the auditory damage that accompanies the ototoxicity of cisplatin is the excessive generation of free radicals although it also has the ability to crosslink DNA and proteins.
Since free radicals may be involved with the toxicity of cisplatin, Lopez-Gonzalez and co-workers (69) examined whether melatonin would also limit the auditory damage caused by this chemotherapy. As in their studies of aminoglycoside toxicity, these workers recorded DPOAEs from the external auditory canal of rats after their treatment with cisplatin alone or in combination with antioxidants. To produce the DPOAEs, the animals received a single intraperitoneal dose of 10 mg/kg cisplatin. Additional animals were given the cancer chemotherapeutic agent in combination with melatonin (10 mg/L in the drinking water or a daily subcutaneous dose of 250 µg) or with a mixture of antioxidants, also injected subcutaneously, that included 0.2 mg alpha-tocopherol, 3 mg ascorbic acid, 1 mg glutathione and 60 mg N-acetylcysteine.
The DPOAEs, recorded as DPgrams, were reduced by both melatonin and the antioxidant mixture to an equal amount on days 0, 5 and 15 after cisplatin administration. The difference, however, was that the antioxidant mixture contained a much higher dose of free radical scavengers than did melatonin alone. As a result, the authors concluded that, in terms of ototoxicity as estimated using DPgrams, melatonin was an estimated 350 times more effective than the antioxidant mixture in preserving the integrity of the auditory apparatus. The importance of these findings is that melatonin interferes with the toxicity of cisplatin, it does not counteract its efficacy as a cancer chemotherapy (70).
Tinnitus is the perception of sound when it is otherwise externally quiet. It occurs as a result of a variety of conditions including allergies, a consequence of medications (e.g., aspirin) or a congenital defect and may accompany natural hearing loss that is observed during aging. Tinnitus is often caused by noise-induced hearing impairment (71). When the health care professional can actually hear a sound emanating from the ear of a patient, it is referred to as objective tinnitus (72). Conversely, where no noise can be perceived by anyone other than the patient, it is referred to as subjective tinnitus (73).
Interest in the potential use of melatonin to minimize tinnitus was initiated by observation of Rosenberg et al. (74) who found that melatonin, taken at a dose of 3 mg daily for 30 days, caused a perceptible reduction in tinnitus. The improvement was most obvious in patients with bilateral, as opposed to unilateral, tinnitus and in those whose sleep was disturbed because of tinnitus. Melatonin is also a sleep promoting agent, however, which may have reduced the perception of tinnitus. Due to their results, the authors suggested that melatonin should be included in the armamentarium of drugs useful in treating subjective tinnitus.
A double-blind, placebo-controlled prospective study also showed that melatonin alleviated subjective tinnitus (75). In this study, melatonin (3 mg daily) was compared with the D2 dopamine receptor antagonist, sulpiride (50 mg daily), or combined with it with tinnitus perception being subjectively graded by the patients; the duration of treatment was 30 days. Using the subjective rating scale, sulpiride reduced tinnitus by 56%, melatonin lowered it by 40% and the combined treatment caused an 81% drop in subjective tinnitus. The visual analogue scale evaluation also indicated that the combination of drugs also was the best treatment to reduce tinnitus perception. In this study it was surmised that melatonins efficacy in limiting the perception of tinnitus stemmed from its ability to reduce the response to dopamine, possibly by inhibiting the release of the tissue biogenic amine which may be mediated by central-type benzodiazepine receptors (76). While Lopez-Gonzalez and co-workers (75) provide a logical flow diagram to illustrate how melatonin may impact tinnitus via a dopaminergic mechanism, the paper provides no direct support for this hypothesis.
Similar findings using a combination of melatonin and sulodexide were observed in terms of reducing tinnitus. In the 34 patients receiving this treatment, the combination of drugs improved the quality of life and reduced the subjective evaluation of the severity of tinnitus (77). Sulodexide, a glycosaminoglycan, was used because of its ability to enhance the blood flow in the labyrinthine microcirculation.
In a study that lacked a placebo-treated control group, melatonin also seemingly reduced the subjective interpretation of the severity of tinnitus in patients as assessed using the Tinnitus Handicap Inventory (78). In the same report, the effect of melatonin on sleep was estimated. As with the presumed reduction in tinnitus, melatonin also improved sleep quality as evaluated using the Pittsburgh Sleep Quality Index. There seemed to be some association between the improvements in sleep and tinnitus. The authors of this report feel melatonin may be a safe treatment for tinnitus, particularly in patients where the tinnitus is so severe it disturbs sleep.
In a recent theoretical report, Pirodda et al. (79) enumerated the means by which melatonin may relieve tinnitus. The proposed mechanisms include melatonins ability, a), to reduce sympathetic drive, b), to induce a more steady hemodynamic condition resulting in a regular labyrinthine perfusion, c), to cause a reduction in skeletal muscle tone which would limit tinnitus that results from tonic contraction of the tensor tympani muscle, d), as a result of its antidepressive actions which may, via indirect means, reduce tinnitus or the perception of tinnitus severity, and, e), as a direct regulator of inner ear immunity.
In general, the findings summarized in the reviewed publications suggest the utility of melatonin in treating tinnitus. Considering how common-place this disturbing condition is, any treatment, especially one with the low cost and high safety profile of melatonin, should be considered for use.
The studies performed to date do not unequivocally establish the mechanisms by which melatonin reduces the toxicity of the aminoglycosides and cisplatin. It is known, however, that the use of these often highly-effective drugs is limited by their negative side effects. If, in fact, melatonin routinely ameliorates their toxicity in humans as it does in animals, then the dose of these drugs could perhaps be increased thereby elevating their effectiveness. To date, there are no studies showing that melatonin, combined with either aminoglycoside or cisplatin treatment, interferes with their action. Rather, the authors of the published reports specifically mention that melatonin does not attenuate the functional efficiency of either the aminoglycosides or cisplatin.
Melatonin, an endogenously-produced molecule, has frequently been used by humans for various purposes (especially as a sleep promoting agent) without evidence of toxicity (80-87). The common doses given to humans usually are in the low milligram range, i.e., 1-10 mg/day. Based on animal studies where melatonin has frequently been used as a pharmacological agent, these low doses in humans may be insufficient to reduce the damage done by toxic drugs like the aminoglycosides and cisplatin. Thus, higher doses should be considered in clinical studies. Melatonin has been given to normal humans at the dose of 1 g daily for 30 days with no untoward side effects being apparent (88).
In view of the observations summarized in the review, it would seem worthwhile to consider combining melatonin with toxic pharmaceuticals for the purpose of reducing the damage these latter molecules often promote at the level of the cochlear and vestibular hair cells. The more wide spread use of melatonin in both the experimental and the clinical setting and in clinical trials of several types has been previously proposed (85, 89).
Conflict of interests: None declared.
- Schacht J. Biochemical basis of aminoglycoside ototoxicity. Otolaryngol Clin North Am 1993; 26: 845-856.
- Rybak LP, Ramkumar V. Ototoxicity. Kidney Int 2007; 72: 931-935.
- Johnsson LG, Hawkins JE Jr, Kingsley TC, Black FO, Matz GJ. Aminoglycoside-induced cochlear pathology in man. Acta Otolaryngol 1981; 381: 1-19.
- Humes DH. Aminoglycoside nephrotoxicity. Kidney Int 1988; 33: 900-911.
- Chen Y, Huang HG, Zha DJ, et al. Aspirin attenuates gentamicin ototoxicity from the laboratory to the clinic. Hear Res 2007; 226: 178-182.
- Guthrie OW. Aminoglycoside-induced ototoxicity. Toxicology 2008; 249: 91-96.
- Minor LB. Gentamicin-induced bilateral vestibular hypofunction. J Am Med Assoc 1998; 279: 541-546.
- Santucci KA, Krieger JN. Gentamicin for the practicing urologist: a review of the efficacy, single daily dosing and switch therapy. J Urol 2000; 163: 1076-1084.
- Nakashima T, Teranichi M, Hibi T, Kobayashi M, Umemura M. Vestibular and cochlear toxicity of aminoglycosides. Acta Otolaryngol 2000; 120: 904-911.
- Davis BD. Mechanisms of bactericidal action of aminoglycosides. Microbiol Rev 1987; 51: 341-350.
- Jezowska-Bojczuk M, Lesniak W. Coordination mode and reactivity of copper (II) complexes with kasugamycin. J Inorg Biochem 2001; 85: 99-105.
- Yang M, Tomellini SA. Non-derivatization approach to high-performance liquid chromatography-fluorescence detection for aminoglycoside antibiotics based on a ligand displacement reaction. J Chromatogr 2001; 939: 59-67.
- Jezowska-Bojczuk M, Karaczyn A, Kozlowski H. Copper (II) binding to tobramycin: potentiometric and spectroscopic studies. Carbohydr Res 1998; 313: 265-269.
- Priuska E, Schacht J. Formation of free radicals by gentamicin and iron: evidence for an iron/gentamicin complex. Biochem Pharmacol 1995; 50: 1749-1752.
- Song BB, Schacht J. Variable efficacy of radical scavengers and iron chelators to attenuate gentamicin ototoxicity in guinea pig in vivo. Hear Res 1996; 94: 87-93.
- Lopez-Gonzalez MA, Delgado F, Lucas M. Aminoglycosides activate oxygen metabolites production in the cochlea of mature and developing rats. Hear Res 1999; 136: 165-168.
- Sha SH, Zajic G, Epstein CI, Schacht, J. Overexpression of copper/zinc-superoxide dismutase protects from kanamycin-induced hearing loss. Audiol Neurootol 2001; 6: 117-123.
- Pirvola IJ, Xing-Oun L, Virkkala J, et al. Rescue of hearing, auditory hair cells, and neurons by CEP-1347/KT7515, an inhibitor of c-Jun N-terminal kinase activation. J Neurosci 2000; 20: 43-50.
- Eshraghi AA, Wang J, Adii E. Blocking c-Jun N-terminal kinase signaling can prevent hearing loss induced by both electrode insertion trauma and neomycin ototoxicity. Hear Res 2007; 226: 168-177.
- Lesniak W, Pecoraro VL, Schacht J. Ternary complexes of gentamicin with iron and lipid catalyze formation of reactive oxygen species. Chem Res Toxicol 2005; 19: 357-364.
- Kennaway DJ, Wright H. Melatonin and circadian rhythms. Curr Top Med Chem 2002; 2: 199-209.
- Reiter RJ. The melatonin rhythm: both a clock and a calendar. Experientia 1993; 49: 654-664.
- Reiter RJ. Pineal melatonin: cell biology of its synthesis and of its physiological interactions. Endocr Rev 1991; 12: 151-180.
- Hardeland R. The presence and function of melatonin and structurally related indoleamines in a dinoflagellate, and a hypothesis on the evolutionary significance of these tryptophan metabolites in unicellulars. Experientia 1993; 49: 614-622.
- Dubbles R, Reiter RJ, Klemke E, et al. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J Pineal Res 1995; 18: 28-31.
- Hattori A, Migitaka H, Iigo M, et al. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptor in vertebrates. Biochem Mol Biol Int 1995; 35: 627-635.
- Paredes SD, Korkmaz A, Manchester LC, Tan DX, Reiter RJ. Phytomelatonin: a review. J Exp Bot 2009; 60: 57-69.
- Tan DX, Hardeland R, Manchester LC, et al. The changing biological roles of melatonin during evolution: from an antioxidant to signals of darkness, sexual selection and fitness. Biol Rev 2010; 85: 607-623.
- Lopez-Gonzalez MA, Guerrero JM, Delgado F. Presence of the pineal hormone melatonin in the rat cochlea: its variation with lighting conditions. Neurosci Lett 1997; 238: 81-83.
- Tan DX, Chen LD, Poeggeler B, Manchester LC, Reiter RJ. Melatonin: a potent, endogenous hydroxyl radical scavenger. Endocrine J 1993; 1: 57-60.
- Rodriguez C, Mayo JC, Sainz RM, et al. Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res 2004; 36: 1-9.
- Tan DX, Manchester LC, Reiter RJ, et al. A novel melatonin metabolite, cyclic 3-hydroxymelatonin: a biomarker of in vivo hydroxyl radical generation. Biochem Biophys Res Commun 1998; 253: 614-620.
- Tan DX, Manchester LC, Burkhardt, et al. N1-acetyl-N2-formyl-5-methoxykynuramine, a biogenic amine and melatonin metabolite, functions as a potent antioxidant. FASEB J 2001; 15: 2294-2296.
- Ressmeyer AR, Mayo JC, Zelosko V, et al. Antioxidant properties of the melatonin metabolite, N1-acetyl-5-methoxykynuramine (AMK): scavenging of free radicals and prevention of protein destruction. Redox Rep 2003; 8: 205-213.
- Hardeland R, Tan DX, Reiter RJ. Kynuramines, metabolites of melatonin and other indoles: the resurrection of an almost forgotten class of biogenic amines. J Pineal Res 2009; 47: 108-126.
- Reiter RJ. Melatonin: lowering the high price of free radicals. News Physiol Sci 2000; 15: 246-250.
- Tan DX, Reiter RJ, Manchester LC, et al. Chemical and physical properties and potential mechanisms: melatonin as a broad spectrum antioxidant and free radical scavenger. Curr Top Med Chem 2002; 2: 181-188.
- Hardeland R. Antioxidative protection by melatonin: multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005; 271: 119-130.
- Konturek PC, Konturek SJ, Celinski K, et al. Role of melatonin in mucosal gastroprotection against aspirin-induced gastric lesions in humans. J Pineal Res 2010; 48: 318-323.
- Barlow-Waldon LR, Reiter RJ, Abe M, et al. Melatonin stimulates brain glutathione peroxidase activity. Neurochem Int 1995; 26: 497-502.
- Pablos MI, Agapito MT, Gutierrez R, et al. Melatonin stimulates the activity of the detoxifying enzyme glutathione peroxidase in several tissues of chicks. J Pineal Res 1995; 19: 111-115.
- Limson J, Nyokong T, Daya S. The interaction of melatonin and its precursors with aluminum, cadmium, copper, iron, lead and zinc: an absorptive voltammetry study. J Pineal Res 1998; 24: 15-21.
- Leon J, Sainz RM, Mayo JC, Tan DX, Reiter RJ. Melatonin and mitochondrial function. Life Sci 2004; 75: 765-790.
- Lopez A, Garcia JA, Escames G, et al. Melatonin protects the mitochondria from oxidative damage reducing oxygen consumption, membrane potential, and superoxide anion production. J Pineal Res 2009; 46: 188-198.
- Paradies G, Petrosillo G, Paradies V, Reiter RJ, Ruggiero FM. Melatonin, cardiolipin and mitochondrial bioenergetics in health and disease. J Pineal Res 2010; 48: 297-310.
- Urata Y, Homna S, Goto S, et al. Melatonin induces gamma-glutamylcysteine synthetase mediated by activator protein-1 in human vascular endothelial cells. Free Rad Biol Med 1999; 27: 838-847.
- Leon J, Escames G, Rodriguez MI, et al. Inhibition of neuronal nitric oxide synthase activity by N1-acetyl-5-methoxykynuramine, a brain metabolite of melatonin. J Neurochem 2006; 98: 2023-2033.
- Reiter RJ, Paredes SD, Manchester LC, Tan DX. Reducing oxidative/nitrosative stress: a newly discovered genre for melatonin. Crit Rev Biochem Mol Biol 2009; 44: 175-200.
- Jou MJ, Peng TI, Yu PZ, et al. Melatonin protects against common deletion of mitochondrial DNA-augmented mitochondrial oxidative stress and apoptosis. J Pineal Res 2007; 43: 389-403.
- Jou MJ, Peng TI, Hsu LF, et al. Visualization of melatonins multiple mitochondrial levels of protection against mitochondrial Ca2+-mediated permeability transition and beyond in rat brain astrocytes. J Pineal Res 2010; 48: 20-38.
- Milczarek R, Hallmann A, Sokolowska K, Kaletha K, Klimek J. Melatonin
enhances antioxidant action of
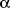 -tocopherol
and ascorbate against NADPH- and iron-dependent lipid peroxidation in human
placental mitochondria. J Pineal Res 2010; 49: 149-155.
-tocopherol
and ascorbate against NADPH- and iron-dependent lipid peroxidation in human
placental mitochondria. J Pineal Res 2010; 49: 149-155.
- Brownell WE. Outer hair cell electromotility and otoacoustic emissions. Ear Hear 1990; 11: 82-92.
- Kemp DT, Ryan S, Bray PA. A guide to the effective use of otoacoustic emissions. Ear Hear 1990; 11: 93-105.
- Henley CM, Owings MH, Stagner BB, Martin GK, Longsbury-Martin BL. Postnatal development of 2f1-f2 otoacoustic emissions in pigmented rat. Hear Res 1989; 43: 141-148.
- Biesalski HK, Welker HA, Thalmann R, Vollrath L. Melatonin and other serotonin derivatives in the guinea pig membraneous cochlea. Neurosci Lett 1988; 91: 41-46.
- Lopez-Gonzalez MA, Guerrero JM, Rojas F, Osuna C, Delgado F. Melatonin and other antioxidants prolong postmortem activity of the outer hair cells of the organ of Corti: its relation to the type of death. J Pineal Res 1999; 27: 73-77.
- Ariano RE, Zelenitsky SA, Kassum DA. Aminoglycoside-induced vestibular injury: maintaining a sense of balance. Ann Pharmacother 2008; 42: 1282-1289.
- Forge A, Schacht J. Aminoglycoside antibiotics. Audiol Neurootol 2000; 5: 3-22.
- Forge A, Li L. Apoptotic death of hair cells in mammalian vestibular sensory epithelia. Hear Res 2000; 39: 97-115.
- Kahlmeter G, Dahlager JI. Aminoglycoside toxicity: a review of clinical studies published between 1975-1982. J Antimicrob Chemother 1984; 13(Suppl A): 9-22.
- Black FO, Peterka RJ, Elardo SM. Vestibular reflex changes following aminoglycoside induced ototoxicity. Laryngoscope 1987; 97: 582-586.
- Song BB, Sha SH, Schacht J. Iron chelators protect from aminoglycoside-induced cochleo- and vestibule-toxicity. Free Rad Biol Med 1998; 25: 189-195.
- Proctor L, Petty B, Lietman P, Glackin R, Shimizu H. A study of potential vestibulotoxic effects of once daily versus thrice daily administration of tobramycin. Laryngoscope 1987; 97: 1443-1449.
- Tran Ba Huy P, Bernard P, Schacht J. Kinetics of gentamicin uptake and release in the rat. Comparison of inner ear tissues and fluids with other organs. J Clin Invest 1999; 77: 1492-1500.
- deVries PJ, Verbooyen RP, Leguit P, Verbrugh HA. Prospective randomized study of once-daily versus thrice-daily netilmicin regimens in patients with intraabdominal infections. Eur J Clin Microbiol Infect Dis 1990; 9: 161-168.
- Kim JB, Jung JY, Ahn JC, Rhee CK, Hwang HJ. Antioxidant and anti-apoptotic effect of melatonin on the vestibular hair cells of rat utricles. Clin Exp Otorhinolaryngol 2009; 2: 6-12.
- Ravi R, Somani SM, Rybak LD. Mechanism of cisplatin ototoxicity: antioxidant system. Pharmacol Toxicol 1995; 76: 386-394.
- Kopke RD, Liu W, Gabaiyadeh R, et al. Use of organotypic cultures of Cortis organ to study the protective effects of antioxidant molecules on cisplatin-induced damage of auditory hair cells. Am J Otol 1997; 18: 559-571.
- Lopez-Gonzalez MA, Guerrero JM, Rojas F, Delgado F. Ototoxicity caused by cisplatin is ameliorated by melatonin and other antioxidants. J Pineal Res 2000; 28: 73-80.
- Reiter RJ, Tan DX, Sainz RM, Mayo JC, Lopez-Burillo S. Melatonin: reducing the toxicity and increasing the efficacy of drugs. J Pharm Pharmacol 2002; 54: 1299-1321.
- Phillips JS, McFerran D. Tinnitus Retraining Therapy (TRT) for tinnitus. Cochrane Database Syst Rev 2010; 17: 3:CD007330.
- Gard AP, Klopper HB, Thorell WE. Successful endovascular treatment of pulsatile tinnitus caused by a sigmoid sinus aneurysm: a case report and review of the literature. Intern Neuroradiol 2009; 15: 425-428.
- Ralli M, Lobarinac E, Fetoni AR, Stolzberg D, Paludetti G, Salvi R. Comparison of salicylate- and quinine-induced tinnitus in rats: development, time course and evaluation of audiologic correlates. Otol Neurotol 2010; 31: 823-831.
- Rosenberg SI, Silverstein H, Rowan PT, Olds MJ. Effect of melatonin on tinnitus. Laryngoscope 1998; 108: 305-310.
- Lopez-Gonzalez MA, Santiago AM, Estaban-Ortego F. Sulpiride and melatonin decrease tinnitus perception modulating the autolimbic dopaminergic pathway. J Otolaryngol 2007; 36: 213-219.
- Tenn CC, Niles LP. Central-type benzodiazepine receptors mediate the antidopaminergic effect of clonazepam and melatonin in 6-hydroxdopamine lesioned rats: involvement of a GABAergic mechanism. J Pharmacol Exp Ther 1995; 274: 84-89.
- Neri G, De Stefano A, Boffa, et al. Treatment of central sensorineural stimulus with orally administered melatonin and sulodexide: personal experience from a randomized controlled study. Acta Otorhinolaryngol Ital 2009; 29: 86-91.
- Megwalu VC, Fennell JE, Piccivillo JF. The effects of melatonin on tinnitus and sleep. Otolaryngol Head Neck Surg 2006; 134: 210-213.
- Pirodda A, Raimondi MC, Ferri GG. Exploring the reasons why melatonin can improve tinnitus. Med Hypotheses 2010; 79: 190-191.
- Simko F, Paulis L. Melatonin as a potential antihypertensive treatment. J Pineal Res 2007; 42: 319-322.
- Aoki K, Zhao K, Yamazaki F, et al. Exogenous melatonin administration modifies cutaneous vasoconstriction response to whole body skin cooling in humans. J Pineal Res 2008; 44: 141-148.
- Robeva R, Kirilov G, Tomova A, Kumanov PH. Melatonin-insulin interactions in patients with metabolic syndrome. J Pineal Res 2008; 44: 52-56.
- Wasdell MB, Jan JE, Bomben MM, et al. A randomized, placebo-controlled study of controlled release melatonin treatment for delayed sleep phase syndrome and impaired sleep maintenance in children with neurodevelopmental disabilities. J Pineal Res 2008; 44: 57-64.
- Gitto E, Pellegrino S, Gitto P, Barberi I, Reiter RJ. Oxidative stress of the newborn in the pre- and postnatal period and the clinical utility of melatonin. J Pineal Res 2009; 46: 128-139.
- Korkmaz A, Reiter RJ, Topal T, Manchester LC, Oter S, Tan DX. Melatonin: an established antioxidant worthy of use in clinical trials. Mol Med 2009; 15: 43-50.
- Chahbouni M, Escames G, Venegas C, et al. Melatonin treatment normalizes plasma pro-inflammatory cytokines and nitrosative/oxidative stress in patients suffering from Duchenne muscular dystrophy. J Pineal Res 2010; 48: 282-289.
- de Castro-Silva C, de Bruin VM, Cunha GM, Nunes DM, Medeiros CA, de Bruin PF. Melatonin improves sleep and reduces nitrite in the exhaled breath condensate in optic fibrosis - a randomized, double-blind, placebo-controlled study. J Pineal Res 2010; 48: 65-71.
- Nordlund JJ, Lerner AB. The effects of oral melatonin on skin color and on release of pituitary hormones. J Clin Endocrinol Metab 1977; 45: 768-774.
- Sanchez-Barcelo A, Mediavilla MD, Tan DX, Reiter RJ. Clinical uses of melatonin: evaluation of human trials. Curr Med Chem 2010; 17: 2070-2095.