Blood-brain barrier (BBB) permeability is
altered in various pathological states, including ischemic stroke. Apart from
being an important mechanism of secondary brain injury, BBB disruption with
plasma protein extravasation was implied in late consequences of acute cerebral
insults, namely epileptogenesis (1-4). The extent of BBB disruption differs
among individual animal models of focal cerebral ischemia (5-9). Many factors
contribute to this variability, including the choice of species, its vascular
anatomy, capacity of collateral circulation, the duration of ischemia (transitory
or permanent), the means of producing ischemia (large vessel occlusion, focal
vasoconstriction, or microvascular occlusion,
e.g. photothrombosis),
and, possibly, the choice of anesthesia. Because BBB disruption in some models
follows a biphasic course (10), the timing of evaluation becomes a major issue
as well.
Usually, investigators focus on BBB disruption in the perilesional tissue. However, focal ischemia was shown to induce changes in the cerebral blood flow, brain metabolism and excitability also in remote functionally connected areas - a concept known as diaschisis (11-14). The possibility that diaschisis could include remote changes in BBB permeability was not tested.
In the present study, we evaluated perifocal, as well as contralateral BBB disruption in photochemically induced focal cerebral ischemia in rats. We also tested, whether the choice of anesthesia can influence the results.
MATERIAL AND METHODS
Adult Male Wistar albino rats (weight 200220 g; ANLAB, Czech Republic) at postnatal
day 60 and older were used. The animals were acclimated at least three days
prior to the study and were maintained on a 12 hour light/dark cycle, with food
and water available ad libitum. All experiments were performed in accordance
with guidelines of the Ministry of Health, Czech Republic. Experimental protocols
were approved by the Ethics committee of the 3
rd
Medical School, Charles University, Prague (Authorization No. for animal use:
17659/2007-30, issued on 9/7/2007; for numbers of funding grants see acknowledgements).
Experimental groups and anesthesia
Sixteen rats were randomly assigned in two experimental groups. Pentobarbital (20 mg/kg i.p.; Sigma, Czech Republic) was used for anesthesia in the first group (PB; N=8), a mixture of ketamine and xylazine (ketamine 80 mg/kg+xylazine 7 mg/kg i.p.; Sigma, Czech Republic) in the latter (KX; N=8). For comparison of background fluorescence signals, a sham-handled control group was created (N=4). Depth of anesthesia was tested at regular 5 min intervals by pressing the skin of the animals hind-limb sole with anatomical tweezers. In case of hind-limb flexion, additional intraperitoneal bolus of half the calculated dose of ketamine was given to ensure surgical anesthesia. Usually, one additional dose of anaesthetic was needed prior to fixation of the animal in the stereotactic frame (approx. 20 minutes after the induction of anesthesia). Throughout the experiment, the animals did not show signs of respiratory distress, or other deficits in vital signs.
Surgery and photothrombosis
Photothrombosis was performed as previously described (15). After the induction
of anesthesia, the scalp of the head was incised (2 cm length in midline) and
the skull overlaying the left sensorimotor cortex was cleaned from soft tissues.
A bolus of photosensitive dye Rose Bengal (RB-Sigma, Czech Republic; 20 mg/2
ml/kg, dissolved in 0.9% NaCl) was applied slowly into the systemic circulation
via tail vein, followed by a bolus of Evans Blue (EB-Sigma, Czech Republic;
0.04 g/kg/2 ml; dissolved in 0.9% NaCl). After the application of dyes, the
animals were positioned in a stereotactic frame. Next, three stereotactically
defined adjacent points on the scull overlaying the hind-limb area of the left
sensorimotor cortex were irradiated by a diode laser beam (532 nm; power density
50 mW/mm
2; illuminated area<1 mm
2).
Each point was irradiated for 6 min (overall 18 min). After the end of photothrombosis,
the animals were left to recover for 24 hours. In contrast to other tracers
(
e.g. natrium fluorescein), EB is almost completely bound to serum albumin.
Thus, EB extravasation is a marker of albumin transport outside the blood vessels.
We chose to apply EB prior to photothrombosis to mirror any change in BBB permeability
within the first 24 hours of ischemia (from the onset of photothrombosis to
brain perfusion and fixation).
Histology, image processing and evaluation
Twenty-four hours after photothrombosis, in deep urethane anesthesia, all animals
were transcardially perfused with a solution of paraformaldehyde and decapitated.
Brains were removed, fixated in paraformaldehyde and sectioned into 40 µm coronary
slices. Lesion dimensions and the distribution of red fluorescence signal emitted
by EB-albumin complex in green light were then studied with a fluorescence microscope
(Olympus
TM). Digital microphotographs of all slices
(both ipsilateral and contralateral cortex) were obtained at fixed image acquisition
parameters (magnification, exposition, sensitivity, resolution, image format).
Additional higher power microphotographs were acquired as needed. All digital
photographs were then analyzed with a freely available utility ImageJ 1.37v
(Wayne Rasband, National Institute of Health, USA;
http://rsb.info.nih.gov/ij/).
1. Ischemic lesions and perifocal blood-brain barrier disruption
Area of necrosis was clearly delimited on each section (
Fig. 3). With
ImageJ tools, the area of necrosis was measured on each slice and multiplied
by the thickness of the section 40 µm. The sum of values from individual slices
gave the volume of necrosis (NV) in an individual animal. The volume of EB-albumin
extravasation (volume of BBB disruption) was measured similarly. The area of
EB-albumin extravasation was delineated manually on each slide. Its border was
defined as the point where clear increase in fluorescence signal was not anymore
observable (
Fig 3A). Again, the sum of values from individual slices
gave the volume of EB-albumin extravasation (EBEV) in an individual animal.
All measurements (608 slices in the PB group, 546 slices in the KX group) were
performed by a single investigator (DK) to avoid inter-rater variability.
2. Remote blood-brain barrier disruption
BBB disruption in the contralateral hemisphere was evaluated by two methods.
Firstly, all slices were visually examined for regions of increased fluorescence
signal and for the occurrence of EB-positive cells (
i.e. cells accumulating
EB-albumin complex). Secondly, we measured differences in background EB-fluorescence
signal between the groups. In every sixth contralateral section, a virtual frame
of 1x2 mm was placed on the contralateral cerebral cortex. The frame was consistently
positioned in a mirror region corresponding to the contralateral area of ischemic
lesion. Mean gray value (mean signal density) was measured within this region
by ImageJ tools (mean gray value= the sum of the gray values of all the pixels
in the selection divided by the number of pixels; in RGB images, each pixel
was converted to gray-scale using the formula: gray=(red+green+blue)/3). Thus,
we have obtained a set of mean gray values (mean signal densities) in all groups
(PB, KX, controls). Differences between the groups were then statistically evaluated.
Statistical evaluation
GraphPad Prism 5.01 (GraphPad Software, Inc.) was used for statistical evaluation of the results. Normal distribution of the results was tested by DAgostino&Pearson omnibus normality test. Statistical significance of differences between PB and KX group with respect to volume of necrosis and volume of EB-albumin extravasation were evaluated by unpaired t-test. Mean signal density values in the contralateral hemisphere were not normally distributed. Therefore, non-parametrical tests (Kruskal-Wallis test, and Mann-Whitney U test) were used to evaluate differences in this parameter among the groups.
RESULTS
Ischemic lesions with central area of necrosis were observed in all animals
of both experimental groups 24 hours after photothrombosis (
Fig. 3A).
As a rule, a large area of EB-albumin extravasation surrounded the lesions,
extending sometimes to the corpus callosum and subcortical structures. In some
slices, brightly red EB-albumin accumulating cells were observed (
Fig. 3B-arrowhead).
Clusters of these cells were usually found at the border of the necrotic core
and the area of BBB breakdown (
Fig. 3B, 3D). EB-stained material was
found in the blood vessels both in the ipsilateral and contralateral hemisphere
and in subcortical structures (see
Fig. 3C, 3D-arrowhead). Pial blood
vessels, as well as perforating blood vessels perpendicular to the pial surface
were sometimes found to be lined with EB-albumin complex. In larger vessels,
EB-albumin complex was found adhering to the vessel wall and escaping into the
extracellular space (
Fig. 3C, 3D-arrowhead).
 |
Fig. 1. Mean volumes of lesions
in groups with different anesthesia. Values are presented as mean ±S.E.M.
Mean volume of necrosis is shown in light gray, mean volume of EB-extravasation
in hatched gray and mean total lesion volume in dark gray. In the group
anesthetized with pentobarbital (n=8), the area of BBB disruption was
significantly larger compared to ketamine-xylazine (n=8) group (p=0.0293,
two-sided unpaired t-test). |
 |
Fig. 2. Median EB-fluorescence
signal density expressed as median gray value in a reference 1x2 mm section
of the hemisphere contralateral to lesion. Median EB-fluorescence signal
density was higher in PB group (p<0.0001, Kruskal-Wallis test with
Dunn’s Multiple Comparison Test). Median EB-fluorescence signal density
in KX group was not different from controls. |
Total volume of lesion (TLV;
i.e. volume of necrosis + volume of EB-albumin
extravasation) did not significantly differ between the groups (TLV
PB=9.4±1.3
mm
3 vs. TLV
KX=8.3±2.1
mm
3). The volume of necrosis (NV) was slightly
larger in the ketamine-xylazine group (NV
PB=5.1±0.7
mm
3 vs. NV
KX=6.3±1.9
mm
3), however, the difference was not statistically
significant. On the other hand, the volume of EB-albumin extravasation (EBEV)
was significantly smaller in the KX group (EBEV
PB=4.3±0.8
mm
3 vs. EBEV
KX=2.0±0.5
mm
3; p=0.0293, two-sided unpaired t-test) (see
Fig. 1).
 |
| Fig.
3. Fluorescence microscopy images of the lesions. (A) Typical
photochemically induced cortical ischemic lesion. Necrotic core can be
seen with a surrounding brightly red area of extravasated EB-albumin.
Lines denote the borders of necrotic core and the area of BBB disruption.
The brightly red material (EB-albumin) in the cortical blood vessels can
be seen in the lower half of the image. (B) A detailed view of
the rim of necrosis and area of BBB failure. EB-albumin accumulating cells
can be seen as bright red dots with a surrounding halo of EB-albumin extravasation
(black arrowheads). (C) A larger blood vessel with EB-albumin
adhering to its walls and escaping into the extracellular space (black
arrowheads). (D) A detail view of the rim of the necrosis with
EB-albumin accumulating cells and a blood vessel extending to the area
of blood brain barrier alteration (black arrowhead). The extravasation
of EB along the blood vessel walls is evident. (E) Low power image
of the hemisphere contralateral to ischemic lesion (animal anesthetized
with PB). A change in the background signal in the left half of the hemisphere
(representing change in blood-brain-barrier permeability) can be noted
(black arrowhead). (F) Signal changes in the medial part of the
hemisphere contralateral to ischemic lesion (animal anesthetized with
PB) (black arrowheads). |
Median EB-fluorescence signal density in the hemisphere contralateral to ischemic
lesion was significantly increased in PB group (p<0.0001, Mann-Whitney U test).
Median EB-fluorescence signal density in the K/X group was similar to the control
group (
Fig. 2). Moreover, in two animals from the pentobarbital group,
diffuse changes in EB-fluorescence signal intensity were found in remote areas
in the contralateral hemisphere (
Fig. 3E, 3F-arrowheads). Unequivocal
EB-albumin accumulating cells were not found in the contralateral cortex.
DISCUSSION
The extent and anatomical pattern of BBB disruption differs among experimental
models of focal ischemia. Breakdown of BBB to proteins is invariably present
in transient middle cerebral artery occlusion (t-MCAO) (16-18), whereas it is
not a typical feature of permanent occlusion model (p-MCAO) (6). Extensive perifocal
vascular leakage is an important feature of photothrombosis (8), and it was
shown to persist at least 24 hours after laser irradiation (19). In agreement
with these findings, we have consistently observed areas of increased EB-albumin
fluorescence surrounding necrotic core of ischemic lesions in the irradiated
area. Areas of increased EB-fluorescence extended into the cerebral white matter
and corpus callosum. Within these regions, we observed adherence of EB-positive
material to the walls of larger penetrating vessels and its leakage into the
extracellular space (
Fig. 3A-3D). Apart from diffuse staining of the
parenchyma surrounding the lesion core, we have observed uptake of EB-albumin
complex into cells (
Fig. 3B, 3D-arrowheads). Plasma protein uptake into
brain cells was described in various models of focal brain ischemia, as well
as other insults (20-25) and may play an important role in epileptogenesis (2-4).
Interestingly, animals subject to photothrombosis (where large BBB breakdown
is typical) frequently develop seizures (26-27).
Our study demonstrates that the extent of BBB disruption in the photothrombotic
model can be significantly reduced when a combination of NMDA antagonist ketamine
and a2-adrenoceptor agonist xylazine is used for anesthesia, in contrast to
GABA
A agonist pentobarbital. Similar results
were observed when selective NMDA antagonist MK-801 was used in the t-MCAO model
(28). We have found no direct evidence in the literature for xylazine-mediated
alteration of blood-brain barrier permeability, although, in one study, a decrease
of ethanol-induced BBB opening was observed following pretreatment with another
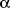
2-adrenoceptor
agonist clonidine (29). On the other hand, the evidence for NMDA receptor mediated
changes to BBB permeability is quite extensive (apart from other important pathophysiological
roles of NMDA receptors,
e.g. in excitotoxicity (30), memory consolidation,
and mood disorders (31). NMDA stimulated uptake of HRP on isolated rat cerebral
cappilaries (32), NMDA antagonist Hu-211 protects against BBB disruption in
photothrombosis (33), intrastriatal injection of NMDA induced extravasation
of Lucifer yellow and this extravasation was prevented by NMDA antagonist (34)
and NMDA applied topically on the cortex increased BBB permeability (35). Nevertheless,
it must be noted, that in some models, this effect of NMDA receptor blockade
on BBB permeability was not reproduced (36).
Although we cannot rule out the role of xylazine in producing the observed effects, we suggest that the presented evidence favors the main role of ketamine in the observed results. The mechanism of NMDA-receptor mediated regulation of BBB permeability remains to be elucidated. Other investigators observed glutamate-induced changes in expression, phosphorylation and distribution of tight-junction proteins, such as occludin (37). Increased transcellular transport of EB-albumin complex in a receptor mediated fashion is also possible (4).
Apart from evaluating perilesional changes in BBB integrity, we also tested
whether focal ischemic lesion can induce BBB alteration in remote (but functionally
connected) areas (diaschisis). The median background EB-fluorescence signal
density was significantly increased in the pentobarbital group (
Fig. 2),
compared to KX and controls. Furthermore, in two animals from the PB group,
we have found regional increase in EB-fluorescence signal density in the medial
part of the contralateral hemisphere (
Fig. 3E, 3F-arrowheads). Also,
in our preliminary experiments in animals anesthetized with PB, we have observed
occasional cellular uptake of EB-albumin in the cerebellum. Although limited,
these findings may support the possibility that the concept of diaschisis is
also relevant to the regulation of permeability of blood-brain barrier and that
alteration of neurotransmission may be involved.
The main limitation of our study is the absence of blood pressure monitoring throughout the experiment. However, with the intraperitoneal route of administration, the risk of hypotension is probably lower than with i.v. application. Moreover, episodes of hypotension would probably tend to influence the results in favor of pentobarbital as in the work by Chi
et al. (38), which was not the case in our study.
In summary, our study shows that anesthesia with NMDA receptor antagonist ketamine decreases the extent of BBB breakdown in cortical photothrombosis. Furthermore, our results indicate that alteration of blood-brain barrier at sites contralateral to photochemically induced ischemic lesion is possible and that it may also be related to the choice of anesthesia. Further studies are needed to broaden these potentially clinically relevant observations.
Acknowledgements:
The work was supported by following grants: Charles Univesity Pregue, research
project UNCE204010; Charles Univesity Pregue, 264706/SVV/2012; Ministry of Education,
Youth and Sports CMS 110/2012.
Conflict of interests: None declared.
REFERENCES
- Cornford EM, Oldendorf WH. Epilepsy and the blood-brain barrier. Adv Neurol 1986; 44: 787-812.
- Seiffert E, Dreier JP, Ivens S, et al. Lasting blood-brain barrier disruption induces epileptic focus in the rat somatosensory cortex. J Neurosci 2004; 24: 7829-7836.
- Bolwig TG. Blood-brain barrier studies with special reference to epileptic seizures. Acta Psychiatr Scand Suppl 1988; 345: 15-20.
- Ivens S, Kaufer D, Flores LP, et al. TGF-beta receptor-mediated albumin uptake into astrocytes is involved in neocortical epileptogenesis. Brain 2007; 130: 535-547.
- Hossmann KA, Olsson Y. The effect of transient cerebral ischemia on the vascular permeability to protein tracers. Acta Neuropathol 1971; 18: 103-112.
- Olsson Y, Crowell RM, Klatzo I. The blood-brain barrier to protein tracers in focal cerebral ischemia and infarction caused by occlusion of the middle cerebral artery. Acta Neuropathol 1971; 18: 89-102.
- Coyle P. Middle cerebral artery occlusion in the young rat. Stroke 1982; 13: 855-859.
- Dietrich WD, Busto R, Watson BD, Scheinberg P, Ginsberg MD. Photochemically induced cerebral infarction. II. Edema and blood-brain barrier disruption. Acta Neuropathol 1987; 72: 326-334.
- Hughes PM, Anthony DC, Ruddin M, et al. Focal lesions in the rat central nervous system induced by endothelin-1. J Neuropathol Exp Neurol 2003; 62: 1276-1286.
- Kuroiwa T, Ting P, Martinez H, Klatzo I. The biphasic opening of the blood-brain barrier to proteins following temporary middle cerebral artery occlusion. Acta Neuropathol 1985; 68: 122-129.
- Hossmann KA, Schuier FJ. Experimental brain infarcts in cats. I. Pathophysiological observations. Stroke 1980; 11: 583-592.
- Kataoka K, Hayakawa T, Yamada K, Mushiroi T, Kuroda R, Mogami H. Neuronal network disturbance after focal ischemia in rats. Stroke 1989; 20: 1226-1235.
- Izumi Y, Haida M, Hata T, Isozumi K, Kurita D, Shinohara Y. Distribution of brain oedema in the contralateral hemisphere after cerebral infarction: repeated MRI measurement in the rat. J Clin Neurosci 2002; 9: 289-293.
- Buchkremer-Ratzmann I, August M, Hagemann G, Witte OW. Electrophysiological transcortical diaschisis after cortical photothrombosis in rat brain. Stroke 1996; 27: 1105-1109, discussion 1109-1111.
- Matejovska I, Bernaskova K, Krysl D, Mares J. Influence of melatonin pretreatment and preconditioning by hypobaric hypoxia on the development of cortical photothrombotic ischemic lesion. Physiol Res 2008; 57: 283-288.
- Yang GY, Betz AL. Reperfusion-induced injury to the blood-brain barrier after middle cerebral artery occlusion in rats. Stroke 1994; 25: 1658-1664, discussion 1664-1665.
- Huang ZG, Xue D, Preston E, Karbalai H, Buchan AM. Biphasic opening of the blood-brain barrier following transient focal ischemia: effects of hypothermia. Can J Neurol Sci 1999; 26: 298-304.
- Nagaraja TN, Keenan KA, Fenstermacher JD, Knight RA. Acute leakage patterns of fluorescent plasma flow markers after transient focal cerebral ischemia suggest large openings in blood-brain barrier. Microcirculation 2008; 15: 1-14.
- Hoff EI, oude Egbrink MG, Heijnen VV, Steinbusch HW, van Oostenbrugge RJ. in vivo visualization of vascular leakage in photochemically induced cortical infarction. J Neurosci Methods 2005; 141: 135-141.
- Loberg EM, Karlsson BR, Torvik A. Neuronal uptake of plasma proteins after transient cerebral ischemia/hypoxia. Immunohistochemical studies on experimental animals and human brains. APMIS 1993; 101: 777-783.
- Loberg EM, Torvik A. Neuronal uptake of plasma proteins in brain contusions. An immunohistochemical study. Acta Neuropathol 1992; 84: 234-237.
- Remmers M, Schmidt-Kastner R, Belayev L, Lin B, Busto R, Ginsberg MD. Protein extravasation and cellular uptake after high-dose human-albumin treatment of transient focal cerebral ischemia in rats. Brain Res 1999; 827: 237-242.
- Sokrab TE, Kalimo H, Johansson BB. Parenchymal changes related to plasma protein extravasation in experimental seizures. Epilepsia 1990; 31: 1-8.
- Matz PG, Lewen A, Chan PH. Neuronal, but not microglial, accumulation of extravasated serum proteins after intracerebral hemolysate exposure is accompanied by cytochrome c release and DNA fragmentation. J Cereb Blood Flow Metab 2001; 21: 921-928.
- Murakami K, Kawase M, Kondo T, Chan PH. Cellular accumulation of extravasated serum protein and DNA fragmentation following vasogenic edema. J Neurotrauma 1998; 15: 825-835.
- Kelly KM, Kharlamov A, Hentosz TM, et al. Photothrombotic brain infarction results in seizure activity in aging Fischer 344 and Sprague Dawley rats. Epilepsy Res 2001; 47: 189-203.
- Kharlamov EA, Jukkola PI, Schmitt KL, Kelly KM. Electrobehavioral characteristics of epileptic rats following photothrombotic brain infarction. Epilepsy Res 2003; 56: 185-203.
- Yang G, Chan PH, Chen SF, Babuna OA, Simon RP, Weinstein PR. Reduction of vasogenic edema and infarction by MK-801 in rats after temporary focal cerebral ischemia. Neurosurgery 1994; 34: 339-345, discussion 345.
- Borisenko SA. Effects of drugs on blood-brain barrier permeability in rats chronically intoxicated by ethanol. Ann Ist Super Sanita 1990; 26: 39-42.
- Wyrembek P, Szczuraszek K, Majewska MD, Mozrzymas JW. Intermingled modulatory and neurotoxic effects of thimerosal and mercuric ions on electrophysiological responses to GABA and NMDA in hippocampal neurons. J Physiol Pharmacol 2010; 61: 753-758.
- Lehner M, Wislowska-Stanek A, Skorzewska A, et al. Expression of N-methyl-D-aspartate (R)(GluN2B) - subunits in the brain structures of rats selected for low and high anxiety. J Physiol Pharmacol 2011; 62: 473-482.
- Koenig H, Trout JJ, Goldstone AD, Lu CY. Capillary NMDA receptors regulate blood-brain barrier function and breakdown. Brain Res 1992; 588: 297-303.
- Belayev L, Busto R, Watson BD, Ginsberg MD. Post-ischemic administration of HU-211, a novel non-competitive NMDA antagonist, protects against blood-brain barrier disruption in photochemical cortical infarction in rats: a quantitative study. Brain Res 1995; 702: 266-270.
- Miller RD, Monsul NT, Vender JR, Lehmann JC. NMDA- and endothelin-1-induced increases in blood-brain barrier permeability quantitated with Lucifer yellow. J Neurol Sci 1996; 136: 37-40.
- Chi OZ, Chang Q, Weiss HR. Effects of topical N-methyl-D-aspartate on blood-brain barrier permeability in the cerebral cortex of normotensive and hypertensive rats. Neurol Res 1997; 19: 539-544.
- Preston E, Webster J, Palmer GC. Lack of evidence for direct involvement of NMDA receptors or polyamines in blood-brain barrier injury after cerebral ischemia in rats. Brain Res 1998; 813: 191-194.
- Andras IE, Deli MA, Veszelka S, Hayashi K, Hennig B, Toborek M. The NMDA and AMPA/KA receptors are involved in glutamate-induced alterations of occludin expression and phosphorylation in brain endothelial cells. J Cereb Blood Flow Metab 2007; 27: 1431-1443.
- Chi OZ, Chun TW, Liu X, Weiss HR. The effects of pentobarbital on blood-brain barrier disruption caused by intracarotid injection of hyperosmolar mannitol in rats. Anesth Analg 1998; 86: 1230-1235.