Our previous work has revealed several differences in the mechanisms of superoxide production between human arteries and veins (13, 14). These findings were described using bypass graft conduit vessels as model systems. The differences between studied vessels are important for understanding of vascular pathology, but also because these vessels are widely used for surgical treatment of coronary artery disease (15). Oxidative stress appears to be more pronounced in human saphenous veins than in mammary arteries, which could partially explain better prognosis in arterial revascularisations (13, 14). However, the contribution of oxidative stress to this process is unknown. Limited number of studies have looked at the effects of superoxide dismutase inhibition on oxidative stress parameters in human blood vessels (16, 17). Those studies have found no significant differences in the increases of superoxide production following SOD inhibition in human arteries and veins. These studies were inconclusive as one found greater SOD activity in arteries while another reported lack of difference. However there was an important drawback in both studies, which used 250 µM-500 µM concentrations of lucigenin that can artificially generate superoxide by redox cycling in the presence of tissues and therefore change the potential differences in superoxide dimutase activity in studied vessels.
Accordingly, we aimed to directly measure superoxide dimutases activity in human arterial and venous bypass conduit vessels; compare their protein expression and functional effects of SOD inhibition on superoxide production.
Patients and Blood Vessels
Excess segments of human saphenous vein (HSV) and internal mammary artery (IMA) were obtained from a total of 27 patients undergoing coronary artery bypass graft surgery. We and others have used this approach to study 'paired' artery and vein specimens, from the same patient, in several previous studies (16-18). Vessels were harvested using a no-touch technique, before surgical distension. The segment was immediately transferred to ice cold Krebs-Hensleit buffer, delicately flushed and carefully dissected to remove excess adventitial tissue, using microsurgical instruments. All vessels were collected before topical administration of drugs such as papaverine. The integrity of the endothelium after harvesting was checked by immunohistochemistry for the endothelial cell marker CD31, in preliminary studies. Patient characteristics and risk factor profile was typical for patients undergoing bypass graft surgery. All patients were treated with HMG-CoA reductase inhibitor.
Collection of tissue specimens was approved by the Local Research Ethics Committees and informed consent was obtained.
Superoxide Dismutase activity
Superoxide dismutase activity was measured by inhibition of pyrogallol autooxidation (19). Frozen vessel segments (-70°C) were ground in liquid nitrogen and suspended in 50 mM Tris buffer, pH 8.2. After centrifugation at 10000 g for 5 minutes, equal volumes of supernatants were mixed with 0.9 ml 50 mM Tris buffer pH 8.2 containing 1 mM DTPA, 10 µM catalase and 200 µM pyrogallol. The change of absorbance at 420 nm was measured every 5 sec for 5 minutes in 23°C using Beckman DU 640 BV spectrophotometer equipped with kinetic software package. The SOD activity was expressed as units/ mg protein, one unit determined as the amount of enzyme that inhibited the oxidation of pyrogallol by 50%. In parallel experiments, the activity of MnSOD was estimated after inhibition of CuZnSOD in samples by the addition of 5 mM KCN.
Western blotting
Portions of vascular homogenate were prepared in a homogenisation buffer (HEPES; 20mM; EDTA 0.01mM; protease inhibitors (Roche); PMSF 1mM; Triton X100 - 2% and SDS 0.5%). Protein content was measured using Bio-Rad system and samples were equalized for protein content. 20 µg protein portions of vascular homogenate were separated by SDS-PAGE (120mV) and transferred to nitrocellulose membranes for 4 hours at 4°C using 200 mAmp in a transfer buffer (25 mMTris; 190 mM Glycine; 20% methanol). MnSOD and Cu/Zn SOD were detected using anti-human SOD sheep polyclonal antibodies (Calbiochem, San Diego, CA). Membranes were incubated for 1 hour at room temperature in 1:500 final dilution of primary antibodies. Anti alpha actin antibody (mAb; Sigma, Germany) (1:2500) was used to check for equal protein loading and data were expressed per a-actin band. Goat horseradish peroxidase (HRP) conjugated anti-sheep IgG was used as secondary antibody to detect SODs (Calbiochem; San Diego, CA). Bands were detected using chemiluminescence substrate and developed on photographic membrane.
Vascular superoxide production
Superoxide production was measured by lucigenin-enhanced chemiluminescence (LGCL), using previously described and validated methods (6, 20). Briefly, intact vessel segments were equilibrated in Krebs-HEPES gassed with 95%O2/5%CO2 for 30 minutes at 37°C. Lucigenin-enhanced chemiluminescence from intact vessels was measured in buffer (2 ml) containing low concentration lucigenin (5 µM). In order to address the role of SOD in modulating superoxide release in human saphenous veins and internal mammary arteries, superoxide release was measured in the presence of SOD inhibitor - diethyldithiocarbamate - DETC (100 µM). Superoxide production was expressed as relative light units (RLU)/second/mg vessel dry weight.
Statistical analysis
Results are expressed as means ± SEM with n equal to the number of patients. Statistical comparisons between the two groups were made using Students t-test for independent or dependent samples. p values <0.05 were considered statistically significant.
Superoxide dismutase activity in human arteries and veins
Since the superoxide dismutase (SOD) enzymes regulate superoxide levels in the vascular wall, we next aimed to investigate whether differences in SOD protein levels and activities could account for differences in the superoxide-mediated effects observed in venous and arterial conduit vessels (Fig.1). Experiments comparing Cu/ZnSOD and MnSOD protein levels using Western Blotting in HSV and IMA revealed no differences between these two conduit vessels (Fig.5 A and B). Similarly, both total SOD activity and MnSOD activity, measured in vascular homogenates using inhibition of pyrogallol auto-oxidation, was similar in both saphenous veins and mammary arteries (Fig.5 C).
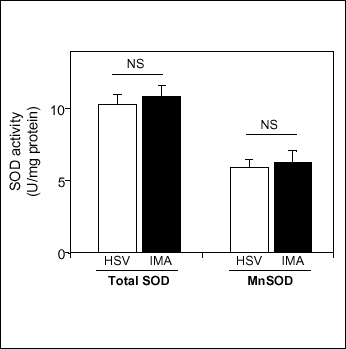 |
Fig. 1. Superoxide dismutase (SOD) activity in human veins and arteries. Total SOD activity was assessed in vascular homogenates in human saphenous vein (HSV) and internal mammary artery (IMA) conduit vessels. Spectrophotometric determination of the inhibition of pyrogallol autooxidation (200 µM) was used. Parallel experiments were conducted in the presence of KCN (5 mM) to inhibit CuZn SOD. Results were calculated in units of activity per mg of total protein. Data are expressed as mean±SEM. NS - lack of significant difference between HSV and IMA. |
SOD protein levels in human venous and arterial conduit vessels
Since the superoxide dismutase (SOD) protein levels may have important effects on SOD activity in the vascular wall, we next aimed to investigate the differences in SOD protein levels between venous and arterial conduit vessels (Fig.2). Experiments comparing Cu/ZnSOD and MnSOD protein levels using Western Blotting in HSV and IMA revealed no differences between these two conduit vessels (Fig.5 A and B).
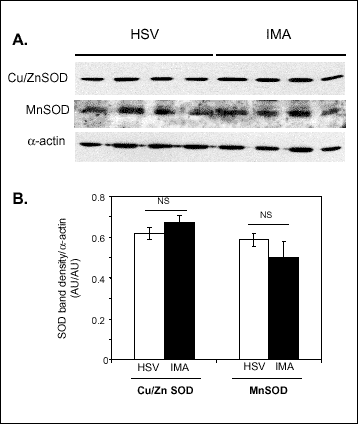 |
Fig. 2. Superoxide dismutase protein levels and activity in venous and arterial bypass conduits Panel A. Western blots showing MnSOD and CuZnSOD proteins in human saphenous vein (HSV) and internal mammary artery (IMA) conduit vessels. alpha-actin band shows equal loading of protein in each sample. Panel B. Densitometric analysis of SOD bands in HSV, IMA (mean±SEM; paired n=6), expressed in relation to density of actin bands. |
Effects of SOD inhibition on vascular superoxide release.
In order to study the functional effect of superoxide dimutase activity in human vessels we measured vascular superoxide production by LGCL in the presence and absence of SOD inhibitor DETC (100 µmol/L). This approach allowed not only to compare the functional effects of vascular SOD in human bypass conduits, but was also an indirect indice of SOD activity measured in intact vessels as opposed to vascular homogenates used by majority of available methods of SOD activity determination.
Vessels were preincubated with DETC for 30 minutes prior to assay. DETC increased ·O2- release in both HSV and in IMA (Fig.3). The degree of increase of superoxide release in response to DETC was similar in HSV and IMA, which was in line with SOD activity measurements.
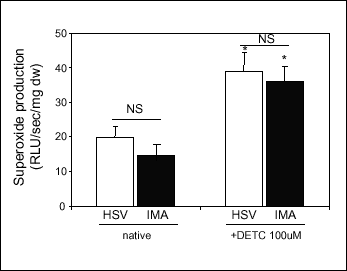 |
Fig. 3. Effects of SOD inhibition by DETC (100 µM) on vascular superoxide release from HSV and IMA (n=8). Superoxide release was determined in intact vascular rings by lucigenin enhanced chemiluminescence (5 µM) in the absence (native) and in the presence of SOD inhibitor - DETC (100 µM). Data were expressed as relative light units per second per mg of dry weight of the vascular ring. Bars indicate mean±SEM. * p<0.05 vs native; NS - lack of significant difference between HSV and IMA. |
Vascular oxidative stress is determined by both production and degradation of free radicals in the vascular wall. While significant amount of data is available about the activity and expression of oxidases in the vascular wall of human bypass conduit vessels, relatively little is known about the anti-oxidant enzyme systems and their role in the regulation of oxidative stress in human veins and arteries.
In the present study, we did not find any significant differences in the activity or expression of superoxide dismutases in the vascular wall of human saphenous veins and internal mammary arteries. This suggests, that the differences in SOD activity are unlikely to be a primary cause for the variable characteristics of oxidative stress in HSV and IMA. Our studies, however, show that in both human saphenous veins and internal mammary arteries, in patients with generalized atherosclerosis, SOD is important in limiting oxidative stress, as its inhibition by DETC caused significant increases in superoxide release from vascular rings. This is in line with studies showing the importance of superoxide dismutase activity in the regulation of NO-dependent endothelial function in both animal models and in humans. In animal models inhibition of SOD by DETC leads to a significant impairment of NO bioavailability (10), and trangenic overexpression of Cu-Zn SOD prevents the development of endothelial dysfunction caused by lipopolysaccharide (21). Landmesser et al. have nicely shown that in patients with coronary artery disease (CAD), vascular EC-SOD activity is substantially reduced. The close relation between endothelium-bound EC-SOD activity and endothelial function suggests that reduced SOD activity contributes to endothelial dysfunction in patients with CAD (22). In our study all vessels were obtained from patients with advanced atherosclerosis undergoing bypass graft surgery. SOD activity could be diminished in this group of patients and the differences in SOD activity could be observed in earlier stages of atherosclerosis or in physiologic conditions. Similarly, activity and expression of superoxide dismutases may differ in younger and elderly individuals (23). Indeed, in young hypercholesterolemic subjects, SOD activity is increased as a defense mechanism to counteract impairment of endothelial function, as the result of increased formation of oxygen free radicals (22). Moreover, although no significant differences in the expression of Cu-Zn SOD or Mn SOD between human conduit artery and vein was found, we cannot exclude a difference in extracellular superoxide dimutase (EC-SOD) protein levels (24), which we were unable to study. Indeed, Landmesser et al. have found that in human coronary arteries from patients with coronary artery disease EC-SOD was increased along with total SOD activity, while Cu-Zn SOD or Mn SOD was not different (22). It is, however, important to note, that both inhibition of pyrogallol autooxidation by vascular homogenates and effects of DETC on superoxide release were not different, indicating that total activity of SOD (which includes EC-SOD) is not different in both types of human vessels.
Some previous studies compared the activity of SOD in human saphenous veins and internal mammary arteries. They have, however, obtained contrasting results. Schmalfuss et al. has found that SOD activity along with the generation of superoxide anion (measured by very high lucigenin concentrations) were all greater in the IMA than in the HSV. Interestingly, Western blot analysis showed no differences in SOD protein expression in IMA and HSV (16). Berry et al., in turn, have found no differences in either activity or protein for SOD (16). Our study brings some additional important insights. Firstly, we used low concentrations of lucigenin in the measurements of superoxide production. Secondly, we paid particular attention to using the same size (weight) vascular rings for both HSV or IMA, which was not discussed by previous studies. Our results of functional effect of SOD inhibition on superoxide release are in agreement with previous studies conducted using high concentrations of lucigenin, thus confirming the validity of the approach taken by Schmalfuss et al. (16) or Berry et al. (17). This may suggest that therapeutic approaches aimed on increasing endogenous antioxidant systems may be very useful in limiting vascular oxidative stress in humans. This approach may be even more important in the light of findings that exogenous antioxidant vitamins do not seem to fulfill this aim. Some novel therapeutic approaches have been proposed recently and could include L-tryptophan (melatonin precursor) which can be useful in re-establishing SOD activity in the model of inflammatory diseases (25). Other potential approaches could include pentoxifylline in inflammation (26), L-carnitine (27), ferulic acid (28) in alcoholic abusive disease, all of which have been shown to induce intrinsic anti-oxidant mechanisms. The mechanisms of SOD activation and regulation of expression are however not fully understood and require further investigation, as novel studies suggest neural component in this process (29).
The determination of mechanisms of oxidative stress in human bypass conduit vessels have important practical implications. Indeed, it has been found that oxidative stress plays an important role in the pathogenesis of intimal hyperplasia in experimental venous bypass conduits (30). Oxidative stress is critical in endothelial dysfunction (2, 3) as well as in the regulation of other factors involved in atherogenesis - including platelet and coagulation regulation (31). The role of superoxide dismutases in this process is not clear. In the present study we show that although the differences in SOD activity can rather not account for differences in the susceptibility to develop graft disease between HSV and IMA, SODs play important limiting role in the oxidative stress pathology in both types of vessels. In previous studies we found that vascular superoxide production has important functional differences between venous and arterial conduits used in patients undergoing coronary artery bypass grafting (4, 14, 18, 20). Specifically, we found that the NAD(P)H oxidase system is quantitatively and proportionately a greater source of superoxide in veins, whereas xanthine oxidase appears to additionally contribute to superoxide production in arteries. We found that increased vascular NAD(P)H oxidase activity is associated with increased protein levels of p22phox, p47phox and p67phox, and increased p22phox and nox2 (gp91phox) mRNA expression (14). The NAD(P)H oxidase is predominantly nox2-based in saphenous veins, whereas a nox4-based oxidase appears proportionately more important in mammary arteries. The importance of differential expression of NAD(P)H oxidase homologues has been shown recently by several investigators (32, 33). Another important difference includes differential response to angiotensin II, which is not observed in veins (17). Despite these differences in the functional and molecular characteristics of superoxide production between veins and arteries, several findings suggest that oxidative stress and endothelial dysfunction are regulated by similar systemic factors in human atherosclerosis. First, both superoxide production and endothelial dysfunction are significantly correlated in arteries and veins from individual patients (14). Second, the expression of both p22phox and nox4 mRNA are strikingly correlated in arteries and veins (14). Finally, protein kinase C signaling has a key role in regulating superoxide production in both veins and arteries (14). The present study adds to that, the superoxide dismutase activity is also a common feature of oxidative stress regulation in veins and arteries. It is also important to remember that systemic nature of oxidative stress may be greatly affected by both clinical risk factors for atherosclerosis including hypertension, hypecholesterolemia or diabetes (18, 20) as well as by genetic factors. The latter may include NAD(P)H oxidase subunit genetic polymorphisms (6), as well as polymorphisms within genes encoding SOD (34).
In conclusion, our study shows that although superoxide dismutase activity and expression do not seem to differ significantly between human conduit arteries and veins, SOD does play a regulatory role in oxidative stress, therefore novel antioxidant approaches are necessary to enhance SOD activity, and improve endothelial function. Some natural products which may be useful in limiting oxidative stress include flavonoids (35) or grapefruit seeds extract (36), as these have been suggested to increase SOD activity in various tissues. However, better understanding of the regulation of vascular antioxidants is necessary to introduce better antioxidant therapeutic strategies for atherosclerosis and graft failure.
- Guzik TJ, Korbut R, Adamek-Guzik T. Nitric oxide and superoxide in inflammation and immune regulation. J Physiol Pharmacol 2003; 54: 469-487.
- Griendling KK, Harrison DG. Dual role of reactive oxygen species in vascular growth. Circ Res 1999; 85: 562-563.
- Harrison DG. Cellular and molecular mechanisms of endothelial dysfunction. J Clin Invest 1998; 100: 2153-2157.
- Channon KM, Guzik TJ. Mechanisms of superoxide production in human blood vessels: relationship to endothelial dysfunction, clinical and genetic risk factors. J Physiol Pharmacol 2002; 53: 515-524.
- Faraci FM, Didion SP. Vascular protection: superoxide dismutase isoforms in the vessel wall. Arterioscler Thromb Vasc Biol 2004; 24: 1367-1373.
- Guzik TJ, West NEJ, Black E, McDonald D, Ratnatunga C, Pillai R, Channon KM. Functional effect of the C242T polymorphism in the NAD(P)H oxidase p22phox subunit on vascular superoxide production in atherosclerosis. Circulation 2000; 102: 1744-1747.
- Mueller CF, Laude K, McNally JS, Harrison DG. Redox mechanisms in blood vessels. Arterioscler Thromb Vasc Biol 2005; 25: 274-278.
- Maczewski M, Duda M, Pawlak W, Beresewicz A. Endothelial protection from reperfusion injury by ischemic preconditioning and diazoxide involves a SOD-like anti-O2- mechanism. J Physiol Pharmacol 2004; 55: 537-550.
- Kukan M. Emerging roles of proteasomes in ischemia-reperfusion injury of organs. J Physiol Pharmacol 2004; 55: 3-15.
- Mugge A, Elwell JH, Peterson TE, Harrison DG. Release of intact endothelium-derived relaxing factor depends on endothelial superoxide dismutase activity. Am J Physiol 1991; 260: C219-C225.
- Didion SP, Ryan MJ, Didion LA, Fegan PE, Sigmund CD, Faraci FM. Increased superoxide and vascular dysfunction in CuZnSOD-deficient mice. Circ Res 2002; 91: 938-944.
- Jiang F, Guo Y, Salvemini D, Dusting GJ. Superoxide dismutase mimetic M40403 improves endothelial function in apolipoprotein(E)-deficient mice. Br J Pharmacol 2003; 139: 1127-1134.
- Guzik TJ, West N, Pillai R, Taggart D, Channon KM. Nitric oxide modulates superoxide release and peroxynitrite formation in human blood vessels. Hypertension 2002; 39: 1088-1094.
- Guzik TJ, Sadowski J, Kapelak B, Jopek A, Rudzinski P, Pillai R, Korbut R, Channon KM. Systemic regulation of vascular NAD(P)H oxidase activity and nox isoform expression in human arteries and veins. Arterioscler Thromb Vasc Biol 2004; 24: 1614-1620.
- Black EA, Guzik TJ, West NE, Campbell K, Pillai R, Ratnatunga C, Channon KM. Minimally invasive saphenous vein harvesting: effects on endothelial and smooth muscle function. Ann Thorac Surg 2001; 71: 1503-1507.
- Schmalfuss CM, Chen LY, Bott JN, Staples ED, Mehta JL. Superoxide anion generation, superoxide dismutase activity, and nitric oxide release in human internal mammary artery and saphenous vein segments. J Cardiovasc Pharmacol Ther 1999; 4: 249-257.
- Berry C, Hamilton CA, Brosnan MJ, Magill FG, Berg GA, McMurray JJ, Dominiczak AF. Investigation into the sources of superoxide in human blood vessels: angiotensin II increases superoxide production in human internal mammary arteries. Circulation 2000; 101: 2206-2212.
- Guzik TJ, West NEJ, Black E, McDonald D, Ratnatunga C, Pillai R, Channon KM. Vascular superoxide production by NAD(P)H oxidase: association with endothelial dysfunction and clinical risk factors. Circ Res 2000; 86: e85-e90.
- Marklund SL, Bjelle A, Elmqvist LG. Superoxide dismutase isoenzymes of the synovial fluid in rheumatoid arthritis and in reactive arthritides. Ann Rheum Dis 1986; 45: 847-851.
- Guzik TJ, Mussa S, Gastaldi D, Sadowski J, Ratnatunga C, Pillai R, Channon KM. Mechanisms of increased vascular superoxide production in human diabetes mellitus: Role of NAD(P)H oxidase and endothelial nitric oxide synthase. Circulation 2002; 105: 1656-1662.
- Didion SP, Kinzenbaw DA, Fegan PE, Didion LA, Faraci FM. Overexpression of CuZn-SOD prevents lipopolysaccharide-induced endothelial dysfunction. Stroke 2004; 35: 1963-1967.
- Landmesser U, Merten R, Spiekermann S, Buttner K, Drexler H, Hornig B. Vascular extracellular superoxide dismutase activity in patients with coronary artery disease: relation to endothelium-dependent vasodilation. Circulation 2000; 101: 2264-2270.
- Kolesnikova EE, Safronova OS, Serebrovskaya TV. Age-related peculiarities of breathing regulation and antioxidant enzymes under intermittent hypoxic training. J Physiol Pharmacol 2003; 54 Suppl 1: 20-24.
- Fukai T, Folz RJ, Landmesser U, Harrison DG. Extracellular superoxide dismutase and cardiovascular disease. Cardiovasc Res 2002; 55: 239-249.
- Leja-Szpak A, Jaworek J, Tomaszewska R, Nawrot K, Bonior J, Kot M, Palonek M, Stachura J, Czupryna A, Konturek SJ, Pawlik WW. Melatonin precursor; L-tryptophan protects the pancreas from development of acute pancreatitis through the central site of action. J Physiol Pharmacol 2004; 55: 239-254.
- Kwiecien S, Brzozowski T, Konturek PC, Pawlik MW, Pawlik WW, Kwiecien N, Konturek SJ. Gastroprotection by pentoxyfilline against stress-induced gastric damage. Role of lipid peroxidation, antioxidizing enzymes and proinflammatory cytokines. J Physiol Pharmacol 2004; 55: 337-355.
- Derin N, Izgut-Uysal VN, Agac A, Aliciguzel Y, Demir N. L-carnitine protects gastric mucosa by decreasing ischemia-reperfusion induced lipid peroxidation. J Physiol Pharmacol 2004; 55: 595-606.
- Rukkumani R, Aruna K, Varma PS, Menon VP. Influence of ferulic acid on circulatory prooxidant-antioxidant status during alcohol and PUFA induced toxicity. J Physiol Pharmacol 2004; 55: 551-561.
- Kwiecien S, Brzozowski T, Konturek PC, Pawlik MW, Pawlik WW, Kwiecien N, Konturek SJ. The role of reactive oxygen species and capsaicin-sensitive sensory nerves in the pathomechanisms of gastric ulcers induced by stress. J Physiol Pharmacol 2003; 54: 423-437.
- West NEJ, Guzik TJ, Black E, Channon KM. Enhanced superoxide production in experimental venous bypass graft intimal hyperplasia: role of NAD(P)H oxidase. Atherosclerosis, Thrombosis, Vascular Biology 2001; 21: 189-194.
- Pajdak W, Radwan J, Guzik TJ. Cleavage of prothrombin bound in immune complexes results in high thrombin enzymatic activity. J Physiol Pharmacol 2004; 55: 477-484.
- Szocs K, Lassegue B, Sorescu D, Hilenski LL, Valppu L, Couse TL, Wilcox JN, Quinn MT, Lambeth JD, Griendling KK. Upregulation of Nox-based NAD(P)H oxidases in restenosis after carotid injury. Arterioscler Thromb Vasc Biol 2002; 22: 21-27.
- Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP, Valppu L, Quinn MT, Lambeth JD, Vega JD, Taylor WR, Griendling KK. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 2002; 105: 1429-1435.
- Ukkola O, Erkkila PH, Savolainen MJ, Kesaniemi YA. Lack of association between polymorphisms of catalase, copper-zinc superoxide dismutase (SOD), extracellular SOD and endothelial nitric oxide synthase genes and macroangiopathy in patients with type 2 diabetes mellitus. J Intern Med 2001; 249: 451-459.
- Schmitt-Schillig S, Schaffer S, Weber CC, Eckert GP, Muller WE. Flavonoids and the aging brain. J Physiol Pharmacol 2005; 56 Suppl 1: 23-36.
- Dembinski A, Warzecha Z, Konturek SJ, Ceranowicz P, Dembinski M, Pawlik WW, Kusnierz-Cabala B, Naskalski JW. Extract of grapefruit-seed reduces acute pancreatitis induced by ischemia/reperfusion in rats: possible implication of tissue antioxidants. J Physiol Pharmacol 2004; 55: 811-821.