The mechanisms of inflammatory response play a crucial role in the pathology of many diseases, including inflammatory bowel disease (IBD) (1) and diseases of atherosclerotic origin (2). Leukocyte infiltration into the intestinal or vascular wall is known to be central to their pathogenesis. Cellular influx into these sites is mediated by a chemical gradient of released chemotactic compounds and an expression of adhesion molecules on endothelial and inflammatory cells (3, 4). The most extensively studied endothelial adhesion molecules include intracellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1), both belonging to immunoglobulins, as well as two selectins: E-selectin and P-selectin. Interactions between endothelial selectins and their ligands located on leukocytes mediate the temporary adhesion and rolling of leukocytes on endothelial cells, while those occurring between ICAM-1 and VCAM-1 and their leukocyte ligands play a key role both in the temporary and permanent adhesion of leukocytes, the latter being dependent on leukocyte activation. Upregulation of endothelial cell adhesion molecules, particularly ICAM-1, in the gut epithelial and vascular tissue is implicated in the pathogenesis of inflammatory bowel disease (5, 6). Furthermore, an increased expression of endothelial adhesion molecules was also observed in atherosclerotic plaques (7), while the levels of circulating soluble form of VCAM-1 were shown to correlate with cardiovascular disease (8).
Most adhesion molecules described above are not present on resting endothelial
cells. Their expression is a result of stimulation by specific molecules, such
as tumor necrosis factor-alpha (TNF-
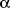
),
interleukin-1 (IL-1), interferon-gamma (INF-
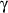
)
and oxidatively modified LDL (oxyLDL) (4, 9) Stimulation is mediated by NF-
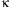
B,
a key transcription factor implicated in the regulation of genes during the
immune and inflammatory responses (10). It has been suggested that peroxisome
proliferator-activated receptor alpha (PPAR
),
a nuclear receptor involved in the transcriptional response to fatty acids,
exerts its anti-inflammatory action through a negative transcriptional regulation
of NF-
B (11). It has been reported that fibric acid derivatives, known to be
PPAR
activators,
inhibit the cytokine-induced expression of VCAM-1 in human endothelial cells
(12).
Fibre is an important nutritional component of the diet. Dietary fibre is a mixture of non-starch polysaccharides from undigested plant cell wall, which serve as substrates for anaerobic fermentation by bacteria in the colon. The majority of cellulose fibres present in the diet are broken down in the large intestine to short chain fatty acids (SCFAs), mainly acetatic, propionaic and butyric acid, of which primarily the former two are being absorbed into the blood (13, 14). High intake of dietary fibre has been associated with an improved bowel function (15). Most of the effects of fibre on gastrointestinal cell function stem from the generation of SCFAs. Harig
et al. (16), for instance, found that SCFAs decreased colonic inflammation in patients with colitis.
Many studies have showed a protective effect of dietary fibre on the outcomes of cardiovascular disease (CVD) (17-20). However, the mechanisms underlying the fibre's influence on the cardiovascular system have yet to be fully elucidated. Recent studies have suggested that the observed action on inflammatory response mechanisms may be of significance for the beneficial effects of high-fibre diet in CVD (20).
Butyrate is, by far, the most extensively studied SCFA, and its anti-inflammatory properties are well documented (21, 22). However, there is little information on acetate and propionate in this respect, although a report published recently has shown that propionic, but not acetic acid accelerated the healing of TNBS-induced colitis (23).
Since it has been previously shown that mononuclear cell recruitment plays a crucial role in the pathogenesis of inflammation and atherosclerosis, we decided to investigate 1) whether propionate inhibits the cytokine-induced expression of VCAM-1 and ICAM-1 and - in consequence - also the monocyte adhesion to endothelial cells, and 2) whether NF- kB and PPAR
play a role in the interactions between cytokines and propionate.
MATERIALS AND METHODS
Materials
Medium 199, fungizone, penicillin, streptomycin, gentamycin, heat-inactivated
bovine serum (FBS), collagenase, gelatine, glutamine, trypsin/EDTA solution
and propionic acid were obtained from Sigma (USA). Recombinant TNF-
(10 mg, 1 x 10
8 U/mg) and IL-1ß (5 x 10
7
U/mg), random hexamers, reverse transcriptase, PCR buffer, Tag polymerase were
purchased from Boehringer Mannheim (Germany). Tissue culture dishes were supplied
by Costar (USA) and Ficoll-Pque by Pharmacia (Sweden). Monoclonal antibodies
against: VCAM-1 (CD 106; FITC), ICAM-1 (CD 54: FITC), CD14 (CD14:PE), CD 45
(CD45:FITC), platelet/endothelial cell adhesion molecule-1 (PECAM-1) (CD31:PE)
and IgG1 (IgG1:FITC) were from Becton Dickinson (USA). Nucleotide dATP, dTTP,
dCTP and dGTP as well as RNAsin were from Promega (USA). ELISA-based kit for
measuring the NF-
B
activation was obtained from Active Motif (Belgium).
Endothelial cell isolation and culture
The experimental protocols were approved by the local Ethic Committee.
Human umbilical vein endothelial cells (HUVEC) were obtained from umbilical
cords by collagenase digestion as described by Jaffe (24). In brief, veins of
umbilical cords were perfused with PBS to remove blood cells, filled with 0.1%
collagenase (type Ia) and incubated for 8 min at 37 °C. The resulting cellular
suspension was supplemented with FBS and centrifuged (300 x g for 10 min). HUVEC
were cultured in gelatine-coated 25 cm
2 flask
and on 6-well or 24-well tissue culture plates, under humidified 5% CO
2
in room air at 37 °C, in Medium 199 supplemented with Earle's salts, 20 mM HEPES,
100 µU/ml penicillin, 100 µg/ml streptomycin, 2.5 µg/ml fungisone, 2 mM glutamine
and 20% FBS. The medium was changed every 2 days up to obtaining confluence
(3-5 days). HUVEC purity was confirmed by the cobblestone morphology, typical
for quiescent endothelial cells, and by PECAM-1 (CD31) staining.
All cells used in this study were of first o second passage.
Peripheral blood mononuclear cells (PBMC) preparation
Heparinized blood was diluted with PBS and 25 ml was immediately layered over 15 ml Ficoll-Paque and centrifuged (400 x g, 40 min, 22 °C). The mixed mononuclear cell band was removed by aspiration, washed with PBS, centrifuged and suspended in HUVEC culture medium with the FBS concentration reduced to 5%. The suspended cells were counted and used in the adhesion assay. PBMC consisted of approximately of 10% monocytes and 90% lymphocytes.
Measurement of VCAM-1 and ICAM-1 expression in HUVEC
Second passage HUVEC were cultured in gelatine-coated 24-well plates. Upon confluence,
FBS concentration was reduced to 5% and cultured without or with propionate
at different concentrations (0.1, 1.0 or 10.0 mM - prepared from a stock solution
of 100 mM by neutralization with 1 M NaOH) for 30 min, 12 h or 24 h. Next, HUVEC
were incubated for 8 h with TNF-
(100 U/ml) or IL-1ß (100 U/ml) in the absence or presence of propionate.
Cell morphology and
viability were examined by measuring lactate dehydrogenase
activity and trypan blue dye exclusion, and were found to remain unchanged under
experimental condition.
Following stimulation, cells were washed with PBS and treated for 30 min at
4 °C with a saturating amount of PE-conjugated anti-PECAM-1 (CD31) and FITC-conjugated
either anti-ICAM-1 (CD54) or anti-VCAM-1 (CD106) monoclonal antibody. For isotype
control, FITC-conjugated mouse anti-IgG
1 was
used. Next, cells were washed with PBS, fixed in 1% paraformaldehyde, harvested
by mild trypsinization, washed again and centrifuged at 300 x g for 10 min.
After that, cells were analysed (10,000 cells per sample) by flow cytometry
(FACS Calibur, Becton Dickinson). After correction for unspecific binding (isotype
control), specific mean fluorescence intensity (MFI) was measured in each channel.
Adhesion assay
HUVEC were grown to confluence on 24-well plates, pretreated with propionate
(10 mM) for 24 h and stimulated with TNF-
(100 U/ml) or IL-1ß (100 U/ml) for 8 h, then the adhesion assay was performed.
Briefly, HUVEC were washed with PBS and co-incubated for 30 min with a PBMC
suspension (1 x 10
6/ml). The PBMC suspension was
removed and the cells were washed twice with PBS to remove nonadherent cells.
Next, the cells consisting of endothelial cells, monocytes and lymphocytes were
incubated for 30 min at 4 °C with saturating amounts of FITC-conjugated mouse
anti-CD45 and PE-conjugated anti-CD14. Following that, cells were washed with
PBS, fixed in 1% paraformaldehyde, washed again, harvested by mild tripsinization,
centrifuged and counted. Cells prepared in this manner were analysed (10 000
cells per sample) by flow cytometry. The proportion of monocytes and lymphocytes
in the suspension was determined by measuring fluorescence I (FLI - CD45) and
fluorescence II (FL II - CD14). The absolute number of monocytes and lymphocytes
adhering to endothelial cells was calculated in relation to the total number
of cells obtained after trypsinization. The results were expressed as a percentage
of monocyte and lymphocyte added.
Quantitation of VCAM-, ICAM-1 and PPAR gene expression
For the quantititation of VCAM-1 and ICAM-1 gene expression, HUVEC were grown
to confluence on 6-well plates, pretreated with propionate (10 mM) for 24 h
and stimulated with TNF-
(100 U/ml) for 4 h. For PPAR
gene quantitation, HUVEC were grown to confluence in 6-well plates and than
incubated with propionate (0.1, 1.0 or 10.0 mM) for 24h. Total RNA (10
6
cells) was isolated by the method of Chomczynski (25) using TRIZOL, and its
concentration was calculated from spectrophotometric measurements at 260 nm.
RNA (500 ng) was dissolved in 20 µl of the reaction mixture containing 2.5 mM
of dATP, dTTP, dCTP, dGTP, 20 U of RNAsin, 100 pM of random hexamers and 20
U of MMLV Reverse Transcriptase. Incubation was carried out at 37 °C for 60
minutes, the temperature of the reaction was then raised to 94 °C for 5 minutes
to inactivate the enzyme and finally reduced to 4 °C. An aliquot of cDNA (5
µl of RT mixture) was dissolved in 25 µl of the reaction mixture containing
10 x PCR buffer (final Mg
2+ concentration: 1.5
mM), 2.5 mM of dATP, dTTP, dCTP, dGTP, 10 pM of up- and downstream primers (PPAR
,
VCAM-1, ICAM-1, GAPDH and ß-actin) and 1 U
Taq polymerase. For
semiquantitative analysis, linearity of amplification of PPAR
,
VCAM-1, ICAM-1, GAPDH and ß-actin cDNAs depending on the PCR cycle number
was established in preliminary experiments. 28 cycles for PPAR
,
16 for VCAM-1, 18 for ICAM-1, 22 for GAPDH and for ß-actin yielded the
best amplification profile to allow for distinguishing the differences among
the samples. The following sets of primers were used in PCR amplification:
VCAM -1: sense 5'`-CCCTTGACCGGCTGGAGATT-3`
antisense 5`-CTGGGGGCAACATTGACATAAAGTG-3`
ICAM-1: sense: 5`-TGAAGGCCACCCCAGAGGACAAC-3`
antisense 5`-CCCATTATGACTGCGGCTGCTGCTACC-3`
GAPDH : sense 5`-GAGTCAACGGATTTGGTCGT-3`
antisense 5`-GTTGTCATGGATGACCTTGG-3`
PPAR
: sense 5`-GCC CCT CCT CGG TGA CTT ATC-3`
antisense 5`-ATG ACC CGG GCT TTG ACC TT-3`
b-actin: sense 5'-CCTCGCCTTTGCCGATCC-3'
antisense 5'GGATCTTCATGAGGTAGTCAGTC-3'
Amplification products obtained in PCR (241 bp for VCAM-1, 409 bp for ICAM-1
482 bp for GAPDH, 454 bp for PPAR
and ß-actin) were separated by SDS-PAGE on a 3% agarose gel. Images of
ethidium bromide-stained bands for PPAR
,
VCAM-1, ICAM-1 and GAPDH were taken with a DS-34 Polaroid camera. The intensity
of the bands was measured by densitometry using the gel analysis macro supplied
with NIH Image software. The signals were normalized (VCAM-1 and ICAM-1 to the
cDNA levels of the GAPDH housekeeping gene, PPAR
to ß-actin) and expressed as a ratio.
Measurement of the NF-B activation
To measure the NF-
B
activation, confluent HUVEC were preincubated with or without propionate (10
mM) for 24 h, followed by exposure to TNF-
(100 U/ml) for 2 h. NF-
B
activation was measured in a whole cell extract using the ELISA-based kit. Briefly,
to obtain the whole cell extract, HUVEC were washed with ice-cold PBS, scraped
into tubes and centrifuged. The pellet was lysed with the use of the Complete
Lysis Buffer containing dithiothreitol and a protease inhibitor cocktail. After
centrifugation at 14 000 rpm at 4 C for 20 min, the protein concentration in
the supernatant (whole cell extract) was determined by using the Bradford protein
assay. To determine the NF-
B
activation with the use of the ELISA-based kit, 10 µg of the whole cell extract
was added to a 96-well plate on which an oligonucleotide containing the NF-
B
consensus binding site (5'- GGGACTTTCC-3') has been immobilized. The active
form of NF-
B contained
in the cell extract specifically bound to this oligonucleotide. After 1 h incubation,
the plate was washed three times and the primary antibody against the active
form of NF-
B was
added for the next 1 h. The antibody recognizes an epitope on p65 that is accessible
only when NF-
B
is activated and bound to its target DNA. After washing, secondary antibody
conjugated to horseradish peroxidase was added to obtain a sensitive readout
by spectrophotometry at 450 nm. The experiment was performed in duplicates and
results are expressed as OD
450 nm.
Statistical analysis
Results are expressed as mean ± SD. Differences were analysed by 1-way ANOVA
followed by Fisher's protected least significant difference test. A value of
p < 0.05 was regarded as significant.
RESULTS
Effects of propionate on VCAM-a and ICAM-1 expression by TNF--stimulated HUVEC
VCAM-1 and ICAM-1 expression on the surface of endothelial cells was evaluated by flow cytometry. Expression was related to the mean fluorescence intensity from FCS (forward scatter) and PECAM-1 (CD 31) gated cells, representing 80-90% of the total pool of cells analysed. Expression of PECAM-1 was regarded as a marker of endothelial
viability, in the light of the fact that it is always present on
viable and disappears from dead cells (30). When compared with control cells, no significant changes were found in the PECAM-1 expression on cells exposed to propionate and TNF-
or IL-1.
As expected, HUVEC cultured for 8 h with TNF-
or IL-1ß demonstrated a marked increase in VCAM-1 and ICAM-1 expression
as compared to resting cells (
Table 1). Pretreatment of HUVEC with propionate
(10 mM) for 24 h significantly reduced both VCAM-1 and ICAM-1 expression levels
in TNF-
-stimulated
cells: by 55-65% (
p < 0.001, n = 7) and 22-32% (
p < 0.001, n =
7), respectively. Expression of VCAM-1 and ICAM-1 on HUVEC stimulated with IL-1ß
was reduced by 37-63% (
p < 0,001, n = 7) for VCAM-1 and by 21-40% (
p
< 0,001, n = 7) for ICAM-1. It appeared that the effect of propionate on TNF-
-induced
VCAM-1 expression was more pronounced as compared to ICAM-1 expression. Treatment
of unstimulated HUVEC with propionate did not alter VCAM-1 and ICAM-1 expression
(data not shown).
| Table 1. Propionate
inhibits TNF--
and IL-1-induced VCAM-1 and ICAM-1 expression in HUVEC. |
 |
| Cells were preincubated
with or without propionate (10.0 mM) for 24 h prior to incubation without
or with TNF-
(100 U/ml) or IL-1 (100 U/ml) for 8 h. Next, cells were stained for VCAM-1
and ICAM-1 and analyzed by flow cytometry. Results are expressed as percent
of TNF-- or
IL-1-stimulated cells (% control) and presented as mean ± SD from 7 experiments.
*p < 0.001 - as compared to cells stimulated with TNF-
or IL-1 in the absence of propionate. |
To investigate the time and concentration dependency of propionate treatment
on the VCAM-1 and ICAM-1 expression, HUVEC were pretreated with propionate at
different concentrations or for different periods of time prior to stimulation
with TNF-
. Inhibition
of the TNF-
-induced
VCAM-1 and ICAM-1 expression depended on propionate concentration, with a maximum
reduction observed at 10 mM (
Fig. 1A). In addition, propionate inhibited
VCAM-1 and ICAM-1 expression induced by TNF-
in a time-dependent manner, with the maximum reduction occurring after 24h of
preincubation with propionate (
Fig. 1B).
 |
Fig. 1. Propionate inhibits
TNF--induced
cell surface expression of VCAM-1 and ICAM-1 in HUVEC. A - Cells were
preincubated with or without propionate (0.1, 1.0, 10.0 mM) for 24 h prior
to incubation without or with TNF-
(100 U/ml) for 8 h. B - Cells were preincubated with or without propionate
(10.0 mM) for 30 min, 12 h or 24 h and then stimulated with TNF-
(100 U/ml) for 8 h. Cells were next stained for VCAM-1 and ICAM-1 and
analyzed by flow cytometry. Results are expressed as percent of TNF--stimulated
cells (% control), and are given as mean ± SD from 7 experiments. #p
< 0.00` as compared to control; ***p < 0.05, **p < 0.01,
*p < 0.001 - as compared to cells stimulated with TNF-
in the absence of propionate. |
Effects of propionate on adhesive properties of TNF-- or IL-1 -stimulated HUVEC towards monocytes and lymphocytes
In an attempt to determine the functional importance of inhibitory properties
of propionate on the TNF-
-
or IL-1ß-induced expression of VCAM-1 and ICAM-1, we have studied the
adhesion of freshly isolated monocytes and lymphocytes to HUVEC. Stimulation
of HUVEC by TNF-
increased the number of adherent monocytes from 19.0% ± 2.9% to 26.3% ± 3.5%
of added monocytes (
Fig. 2A) and the number of lymphocytes from 2.0%
± 0.7% to 11.2% ± 1.5% of delivered lumphocytes (
Fig. 2B). The same pattern
was observed when TNF-
was replaced by IL-1ß. The number of adherent monocytes increased from
19.0% ± 2.0% to 25.11% ± 3.7% (
Fig. 2A) and lymphocytes from 2.0% ± 0.5%
to 10.3% ± 1.4% (
Fig. 2B) of added cells. Pretreatment with 10 mM propionate
for 24 h significantly reduced (
p < 0.01, n = 9) the adhesion of monocytes
(
Fig. 2A) to HUVEC from 26.3% ± 3.5% to 19.6% ± 6.0% of cells added to
TNF-
-stimulated
HUVEC, and from 25.11% ± 3.7% to 19.11% ± 5.2% of cells added to IL-1ß-stimulated
HUVEC. The extent of reduction varied in individual populations, but the resultant
mean value of adherent monocytes was similar to unstimulated cells. The results
show that propionate completely abolished the TNF-
-
or IL-1ß-induced increase of HUVEC adhesive properties towards monocytes.
The adhesion of lymphocytes to TNF-
-stimulated
HUVEC was reduced from 11.2% ± 1.5% to 7.5% ± 1.9% of added cells (
p
< 0.001, n = 9) and to IL-1ß-stimulated HUVEC from 10.3% ± 1.4% to 6.6%
± 1.7% of added cells (
p < 0.001, n =9) (
Fig. 2B).
 |
Fig. 2. Propionate inhibits
adhesion of monocytes (A) and lymphocytes (B) to TNF--stimulated
HUVEC.
Endothelial cells were preincubated with or without propionate (10.0 mM)
for 24 h prior to incubation without or with TNF-
(100 U/ml) or IL-1 (100 U/ml) for 8 h. Freshly isolated PBMC were coincubated
with these cells for 30 min. Next, monocyte and lymphocyte adhesion was
measured as described in Materials and Methods. Results are expressed
as a percentage of monocyte and lymphocyte added, and are given as mean
± SD from 9 experiments. #p < 0.001
as compared to control; **p < 0.01, *p < 0.001 - as compared
to cells stimulated with TNF-
in the absence of propionate. |
Effects of propionate on mRNA levels of VCAM-1 and ICAM-1 in HUVEC stimulated by TNF-
To explore the mechanism responsible for inhibition of VCAM-1 and ICAM-1 surface
expression, we studied these molecules' mRNA levels using RT PCR. Stimulation
of HUVEC with TNF-
for 4 h resulted in an approximately 13-fold increase in the amount of specific
VCAM-1 PCR products (
Fig. 3A). Pretreatment with propionate (10 mM) for
24 h significantly reduced the induction of VCAM-1 transcripts by TNF-
.
The reduction varied in individual populations, ranging from 68% to 80% (from
0.52 ± 0.09 to 0.29 ± 0.09;
p < 0.001, n =5). Stimulation of HUVEC with
TNF-
for 4 h induced
only an approximately 2-fold increase in the amount of specific ICAM-1 PCR products
(
Fig. 3A). Similarly to the observed effect on the protein expression,
the inhibitory action of propionate on the TNF-
-mediated
ICAM-1 mRNA level was less pronounced than that on VCAM-1 mRNA. Preincubation
with propionate reduced the induction of ICAM-1 mRNA by 9-13% (from 0.97 ± 0.07
to 0.91 ± 0.08;
p < 0.05; n =5). Neither TNF-
nor propionate altered GAPDH transcription.
 |
Fig. 3. Propionate inhibits
VCAM-1 (A) and ICAM-1 (B) mRNA expression in HUVEC. Cells were preincubated
with or without propionate (10.0 mM) for 24 h prior to incubation without
or with TNF-
(100 U/ml) for 4 h. mRNA was extracted, reverse transcribed and amplified
by PCR using specific primers for GAPDH, VCAM-1 and ICAM-1. A and B -
Optical density from 5 separate experiments after normalization to GAPDH
as a internal standard. Data are expressed as a ratio VCAM-1/GAPDH, and
are given as mean ± SD from 5 experiments. #p
< 0.001 as compared to control; ***p < 0.05, *p < 0.001
- as compared to cells stimulated with TNF-
in the absence of propionate.
Bottom figures show photographs of a UV-illuminated electrophoretic gel from a representative experiment. |
Effects of propionate on the TNF--induced NF-B p65 activation
To determine the effect of propionate on the transcriptional regulation of adhesion
molecule expression, we studied NF-
B
p65 protein levels in the whole cells extract of TNFa-treated HUVEC by new ELISA-based
assay. TNF-
-stimulated
HUVEC showed a marked increase (about 7-fold) in the NF-
B
p65 levels (
Fig. 4). Pretreatment of HUVEC with 10 mM propionate for
24 h significantly reduced (from 0.44±0.061 to 0.184±0.069;
p < 0.001;
n= 9) the NF-
B
p65 activation as compared to TNF-
treated cells. The extent of reduction varied in individual populations, ranging
from 46% to 76%.
 |
Fig. 4. Propionate inhibits
TNF--induced
activation of NF-B.
Cells were preincubated with or without propionate (10.0 mM) for 24 h
prior to incubation without or with TNF-
(100 U/ml) for 2 h. Next, a whole cell extract was obtained and NF-B
activation was measured with the ELISA-based kit. Results are expressed
as OD450nm, and are given as mean ± SD
from 9 experiments. #p < 0.001 as
compared to control; *p < 0.001 - as compared to cells stimulated
with TNF-
in the absence of propionate. |
Effects of propionate on the PPAR expression
Since it has been previously suggested that PPAR
may negatively interact with the activated NF-
B
signalling pathway, we decided to examine whether propionate is a PPAR
activator. RT PCR analysis revealed that incubation of HUVEC with propionate
(10 mM) for 24 h resulted in a significant increase in the amount of specific
PPAR
PCR products
from 0.7 ± 0.11 to 1.1 ± 0.12 (
p < 0.05; n = 6) (
Fig. 5). The
observed effect varied in individual populations, ranging from 44% to 66%. Propionate
had no effect on the constitutive ß-actin transcription.
 |
Fig. 5. Propionate induces
PPAR mRNA
expression in HUVEC. Cells were incubated with propionate (10.0, 1.0 or
0.1 mM) for 24 h. mRNA was extracted, reverse transcribed and amplified
by PCR using specific primers for ß-actin and PPAR.
Top, optical density from 6 separate experiments after normalization to
ß-actin as a internal standard. Data are expressed as a ratio PPAR/ß-actin,
and are given as mean ± SD from 6 experiments. ***p < 0.05 - as
compared to control cells.
Bottom figures show photographs of a UV-illuminated electrophoretic gel from a representative experiment. |
DISCUSSION
Adhesion and transendothelial migration of leukocytes into surrounding tissue are crucial steps in inflammation, immunity and atherogenesis (2, 4). Vascular endothelial cells play an active role in these processes by expressing cell adhesion molecules. It now appears from our findings that propionate is capable of interfering with these pathologically important processes.
In the present study, we have demonstrated for the first time that propionate inhibits TNF-
or IL-1-induced expression of VCAM-1 and ICAM-1, by reducing both protein and specific mRNA levels through inhibition of NF-
B. This effect was significantly more pronounced for VCAM-1 than for ICAM-1, and was time- and dose-dependent. The observed inhibition was associated with a decreased adhesion of monocytes and lymphocytes to stimulated endothelial cells.
Propionate is a naturally occurring SCFA and its levels may be increased by
a more frequent introduction of fibre-rich products into the diet or consumption
of the so-called functional food containing acidophilic bacteria (13, 14). Along
with two other SCFAs, acetate and butyrate, propionate is produced in the large
intestine in the process of bacterial fermentation of cellulose fibres. To date,
butyrate has been the most extensively studied SCFA of the three. Its anti-inflammatory
properties, stemming from its inhibitory effect on NF-
B
activation, among other things, as well as its clinical benefits in the treatment
of inflammatory diseases of the alimentary tract are well documented (21). Recently,
we have shown that butyrate inhibits the cytokine-induced expression of VCAM-1
and ICAM-1 by acting on NF-
B
(22). The observed effect was associated with a decreased adhesion of monocytes
and lymphocytes to stimulated endothelial cells. The attention paid mainly to
butyrate resulted from the fact that of the three SCFAs produced in the large
intestine, acetic and propionic acid pass into the blood, whereas butyric acid
is almost entirely absorbed by colonic epithelium with only a small portion
of butyrate passing into the blood. It is therefore generally assumed that it
is butyrate that is the main contributor to the benefits of a diet rich in cellulose
fibres for the prophylaxis and treatment of alimentary tract diseases. However,
we have demonstrated, for the first time, that also propionate may attenuate
an ongoing inflammatory response at the cellular level and that it may contribute
to the anti-inflammatory properties of SCFAs mixture observed
in vivo.
Notably, it has been shown recently that sodium propionate administrated orally
significantly accelerated the healing of colitis, while administration of sodium
acetate did not produce a similar effect (23).
Our results also show that the propionate-mediated inhibition of cytokine-induced
ICAM-1 and VCAM-1 protein expression was concomitant with a decrease in the
levels of specific mRNA. Thus, it may be suggested that the observed effects
of propionate may have their origin in its influence on the mechanisms regulating
the transcription of genes encoding these molecules. In order to examine further
this thesis, we decided to investigate the influence of propionate on NF-
B,
the key transcription factor implicated in the regulation of variety of genes
during immune and inflammatory response, including those for VCAM-1 and ICAM-1
(10). Most of NF-
B
present in unstimulated cells is regulated by association with one of the many
inhibitory IkB molecules. It can be activated by diverse signals for example
TNF-
, IL-1 or oxyLDL,
which induce the release of NF-
B
from its complex with I
B.
The release of NF-
B
is followed by a translocation of active p50/p65 dimers to the cell nucleus
and their binding to gene promoter regions. We have shown that the levels of
active form of p65 in endothelial cells are increased by the presence of TNF-
and decreased by a pretreatment with propionate. The amount of data on the influence
of propionate on NF-
B
is limited. Recently, a report has been published demonstrating that propionate,
similarly to butyrate, may inhibit the TNF-
-mediated
activation of NF-
B
in colon adenocarcinoma cell line (26).
In our study, we also show that propionate is an activator of PPAR
in endothelial cells. PPAR
exerts anti-inflammatory effects both in the vascular wall and the liver (11). It has been suggested that the anti-inflammatory properties of PPAR
stem from its inhibitory activity on NF-
B activation. PPAR
may limit the endothelial responses that promote leukocyte adhesion and entry into the vessel wall. It has been previously shown that the inhibition of transcriptional expression of VCAM-1, but not ICAM-1, by a PPAR
agonist results, at least partially, from NF-
B inhibition (12). Thus, the activation of PPAR
by propionate observed in this study could potentially explain the fact that this compound exerted a significantly more pronounced inhibitory effect on VCAM-1 than on ICAM-1. Notably, these findings appear to be of particular relevance for the antiatherogenic properties of propionate. It has been suggested that VCAM-1 plays a significant role in the selective accumulation of monocytes and lymphocytes, but not granulocytes, in the walls of blood vessels in atherosclerosis (27).
Several clinical trials have established an inverse relation between fibre intake and coronary heart disease (CHD) (17-19). A higher intake of dietary fibre was associated with a lower risk of CVD and MI (18), and was implicated in protection against intima-media thickness (19). Functional food containing acidophilic bacteria (ProViva) has been shown to lower the LDL cholesterol and fibrinogen levels and decrease the triglyceride levels and systolic blood pressure in patients with moderately elevated cholesterol levels (28) and in smokers (29). Monocytes freshly isolated from the blood of smokers consuming ProViva exhibited a statistically significantly decreased adhesion both to unstimulated and stimulated endothelial cells. Recently, it has been show that high-fibre diet was associated with a reduction in oxidative stress (30). Moreover, a reduction in sICAM-1, sE-selectin and CRP levels, as well as in MCP-1 production, was observed.
In view of the above findings, it transpires that consumption of high-fibre
diet may significantly influence the course of diseases having their origin
in atherosclerosis. Still, the underlying molecular mechanisms of effects of
such a diet require further investigation. The anti-inflammatory mechanisms
of propionate action observed in this study are most likely implicated in the
beneficial action of high-fibre diet in diseases of the alimentary tract. They
also seem to indicate that propionate may be at least partially responsible
for the antiatherosclerotic effects of high-fibre diet. A recent study has demonstrated
an association between dietary fibre and the levels of CRP, a clinical indicator
of inflammation, suggesting that inflammation may be an important mediator in
the association between dietary fibre and CVD (31). From our study it appears
that the beneficial antiatherosclerotic properties of propionate may also stem
from the fact that this molecule is an activator of PPAR
.
Activation of PPAR
results in a reduction of plasma triglycerides (TG), through an induction of
genes that decrease the ability of TG for hepatic very low-density lipoprotein
(VLDL) secretion and which promote the lipoprotein lipase-mediated lipolysis
of TG-rich plasma lipoproteins (32). The stimulatory effect of PPAR
on the plasma level of HDL appears to be mediated by an increased production
of ApoA1 and ApoA2. Kawase
et al. (33) have recently shown that supplementation
of the diet with fermented milk containing
Lactobacillus casei and
Streptococcus
thermophilus results in significant increase in HDL cholesterol and decrease
in triglyceride levels in healthy volunteers. Trials in hyperlipidemic and diabetic
patients showed that high dietary fibre can reduce the plasma levels of TG and
cholesterol. The effect on TG was more marked than on cholesterol. It has been
suggested that the observed hypotriglyceridemic effect may be due to a lower
hepatic secretion rate of TG, through a selective exposure of the liver to an
increased amount of propionic acid produced in the large intestine during fermentation
of non-digestible carbohydrates. The hypocholesterolemic effect is likely due
to a decreased secretion of VLDL. Activation of PPAR
has been also suggested to improve insulin resistance (32). It has been postulated
that propionic acid may improve glucose tolerance and inhibit cholesterol synthesis
in the liver by improving insulin resistance (35-37). On the other hand, you
should not forget that in the case of chronic use of high-fat diet, overexpression
of PPAR
is possible,
which in turn leads to accumulation of triacylglycerols and ceramides in many
organs, including the heart (38). Since ceramides are lipotoxicity mediators,
this may even result in cardiomyopathy.
The particular utilitarian significance of our studies is associated with the fact that propionate could potentially have a strong inhibitory effect on the inflammatory process stimulating the progression of atherosclerosis. Unfortunately, previous attempts of using synthetic non steroidal anti-inflammatory drugs in clinical practice have failed due to their adverse impact on hypertension and coagulation (38). At the same time, an adequate amount of fiber in diet could constitute a safe source of natural anti-inflammatory factors, including propionate. According to the AHA recommendations, this amount should be 25 g of soluble fiber daily, while in countries of high atherosclerosis risk it usually reaches 50% of the demand at the most (39). Thus, although the best known chemical derivative of propionate the widely used ibuprofen - cannot be recommended for lowering the risk of atherosclerosis, the use of fiber as its alternative seems fully justified. It should be added that not only short-chain fatty acids originating from fibre are able to reduce inflammation but similar molecular mechanisms exist also in the case of plant polyphenols originating from the traditional Mediterranean diet, which is currently commonly recommended in atherosclerosis prophylaxis (41).
In conclusion, it should be noted that propionate inhibits the cytokine-induced expression of VCAM-1 and ICAM-1 and, in consequence, the adhesion of mononuclear leukocytes to the endothelial cells, through inhibition of NF-
B activation. The anti-inflammatory and antiatherosclerotic activity of propionate may be associated with its ability to activate PPAR
. Our findings warrant further investigations into the potential therapeutic usefulness of propionate in several pathological events involving leukocyte recruitment, including vascular disease. With this in mind, it could be of particular interest to consider different means of stimulating the production of propionate such as, for example, by ingestion of high amounts of dietary fibre.
Conflict of interests: None declared.
REFERENCES
- Hanauer SB. Inflammatory bowel disease: epidemiology, pathogenesis, and therapeutic opportunities. Inflamm Bowel Dis 2006; 12 (Suppl 1): S3-S9
- Ross R. Atherosclerosis-an inflammatory disease. N Engl J Med 1999; 340: 115-126.
- Jang Y, Lincoff AM, Plow EF, Topol EJ. Cell adhesion molecules in coronary artery disease. J Am Coll Cardiol 1994; 24: 1591-1601.
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell 1994; 76: 301-314.
- Brannigan AE, Watson RW, Beddy D, Hurley H, Fitzpatrick JM, O'Connell PR. Increased adhesion molecule expression in serosal fibroblasts isolated from patients with inflammatory bowel disease is secondary to inflammation. Ann Surg 2002; 235(4): 507-511.
- Vainer B, Nielsen OH. Changed colonic profile of P-selectin, platelet-endothelial cell adhesion molecule-1 (PECAM-1), intercellular adhesion molecule-1 (ICAM-1), ICAM-2, and ICAM-3 in inflammatory bowel disease. Clin Exp Immunol 2000; 121: 242-247.
- Davies MJ, Gordon JL, Gearing AJ et al. The expression of the adhesion molecules ICAM-1, VCAM-1, PECAM, and E-selectin in human atherosclerosis. J Pathol 1993; 171: 223-229.
- Ruef J, Marz W, Winkelmann BR. Markers for endothelial dysfunction, but not markers for oxidative stress correlate with classical risk factors and the severity of coronary artery disease. (A subgroup analysis from the Ludwigshafen Risk and Cardiovascular Health Study). Scand Cardiovasc J 2006; 40: 274-279.
- Kume N, Cybulsky MI, Gimbrone MA Jr. Lysophosphatidylcholine, a component of atherogenic lipoproteins, induces mononuclear leukocyte adhesion molecules in cultured human and rabbit arterial endothelial cells. J Clin Invest 1992; 90: 1138-1144.
- Collins T. Endothelial nuclear factor-kappa B and the initiation of the atherosclerotic lesion. Lab Invest 1993; 68: 499-508.
- Zandbergen F, Plutzky J. PPARalpha in atherosclerosis and inflammation. Biochim
Biophys Acta 2007; 1771: 972-982.
- Marx N, Sukhova GK, Collins T, Libby P, Plutzky J. PPARalpha activators inhibit
cytokine-induced vascular cell adhesion molecule-1 expression in human endothelial
cells. Circulation 1999; 99: 3125-3131.
- Pomare EW, Branch WJ, Cummings JH. Carbohydrate fermentation in the human colon and its relation to acetate concentrations in venous blood. J Clin Invest 1985; 75: 1448-1454.
- Nordgaard I, Mortensen PB, Langkilde AM. Small intestinal malabsorption and colonic fermentation of resistant starch and resistant peptides to short-chain fatty acids. Nutrition 1995; 11: 129-137.
- Trock B, Lanza E, Greenwald P. Dietary fiber, vegetables, and colon cancer: critical review and meta-analyses of the epidemiologic evidence. J Natl Cancer Inst 1990; 82: 650-661.
- Harig JM, Soergel KH, Komorowski RA, Wood CM. Treatment of diversion colitis with short-chain-fatty acid irrigation. N Engl J Med 1989; 320: 23-28.
- Mozaffarian D, Kumanyika SK, Lemaitre RN, Olson JL, Burke GL, Siscovick DS. Cereal, fruit, and vegetable fiber intake and the risk of cardiovascular disease in elderly individuals. JAMA 2003; 289: 1659-1666.
- Liu S, Buring JE, Sesso HD, Rimm EB, Willett WC, Manson JE. A prospective study of dietary fiber intake and risk of cardiovascular disease among women. J Am Coll Cardiol 2002; 39: 49-56.
- Wu H, Dwyer KM, Fan Z, Shircore A, Fan J, Dwyer JH. Dietary fiber and progression of atherosclerosis: the Los Angeles Atherosclerosis Study. Am J Clin Nutr 2003; 78: 1085-1091.
- King DE. Dietary fiber, inflammation, and cardiovascular disease. Mol Nutr Food Res 2005; 49: 594-600.
- Wachtershauser A, Stein J. Rationale for the luminal provision of butyrate in intestinal diseases. Eur J Nutr 2000; 39: 164-1671.
- Zapolska-Downar D, Siennicka A, Kaczmarczyk M, Kolodziej B, Naruszewicz M.
Butyrate inhibits cytokine-induced VCAM-1 and ICAM-1 expression in cultured
endothelial cells: the role of NF-kappaB and PPARalpha. J Nutr Biochem 2004;
15: 220-228.
- Uchida M, Mogami O. Milk whey culture with Propionibacterium freudenreichii ET-3 is effective on the colitis induced by 2,4,6-trinitrobenzene sulfonic acid in rats. J Pharmacol Sci 2005; 99: 329-334.
- Jaffe, E.A. Biology of Endothelial Cells. Boston, MA., Martinus Niijhoff, 1984.
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 1987; 162: 156-159.
- Tedelind S, Westberg F, Kjerrulf M, Vidal A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: a study with relevance to inflammatory bowel disease. World J Gastroenterol 20078; 13: 2826-2832.
- Huo Y, Ley K. Adhesion molecules and atherogenesis. Acta Physiol Scand 2001; 173: 35-43.
- Bukowska H, Pieczul-Mroz J Jastrzebska M, Chelstowski K, Naruszewicz M. The effect of Lactobacillus plantarum in subjects with moderately elevated cholesterol. Atherosclerosis 1997; 137: 437-438.
- Naruszewicz M, Johansson ML, Zapolska-Downar D, Bukowska H. The effect of lactobacillus Plantarum 299v on cardiovascular risk factor in smokers. Am J Clin Nutr 2002; 76: 1249-12155.
- Roberts CK, Chen AK, Barnard RJ. Effect of a short-term diet and exercise intervention in young on atherosclerotic risk factor. Atherosclerosis 2007; 191: 98-106.
- Ignarro LJ, Balestrieri ML, Napoli C. Nutrition, physical activity and cardiovascular disease. An update. Cardiovasc Res 2007; 73: 326-340.
- Duval C, Muller M, Kersten S. PPARalpha and dyslipidemia. Biochim Biophys
Acta 2007; 177: 961-971.
- Kawase M, Hashimoto H, Hosoda M, Morita H, Hosono A. Effect of administration of fermented milk containing whey concentrate to rats and healthy men on serum lipids and blood pressure. J Dairy Sci 2000; 83: 255-263.
- Beylot M. Effects of inulin-type fructans on lipid metabolism in man and in animal models. Br J Nutr 2005; 93 (Suppl 1): 163-168.
- Jenkins DJ, Leeds AR, Gussall MA, Cochet B, Alberti GM. Decrease in posprandial insulin and glucose concentration by guar and pectin. Ann Intern Med 1997; 86: 20-23.
- Venter CS, Vorster HH. Possible metabolic consequences of fermentation in the colon for humans. Med Hypotheses 1989; 29: 161-166.
- Nishina PM, Freedland RA. Effects of propionate on lipid biosynthesis in isolated rat hepatocytes. J Nutr 1990; 120: 668-673.
- Baranowski M, Blachnio A, Zabielski P, Gorski J. PPARlpha agonist induces the accumulation of ceramide in the heart of rats fed high-fat diet. J Physiol Pharmacol 2007; 58: 57-72.
- Garcia Rodriguez LA, Tacconelli S, Patrignani P. Role of dose potenzy in the prediction of risk of myocardial infarction associated with nonsteroidal anti-inflammatory drugs in the general population. J Am Coll Cardiol 2008; 52: 1628-1636.
- Stone NJ, Van Horn L. Therapeutic lifestyle change and Adult Treatment Panel III: evidence then and now. Curr Atheroscler Rep 2002; 4: 433-443.
- Stalinska K, Guzdek A, Rokicki M, Koj A. Transcription factors as targets of the anti-inflammatory treatment. A cell culture study with extracts from some Mediterranean diet plants. J Physiol Pharmacol 2005; 56 (Suppl 1): 157-169.