Magnocellular neurons of the PVN and SON are the main hypothalamic sources of oxytocin (OXY) and vasopressin (AVP) (1, 2) which play a principal role in the maintenance of body fluid homeostasis (3-5). Besides AVP and OXY, other biologically active substances (6-8), including TH (9-13), CRH (14), and NPY (15), have been identified in the magnocellular neurons. Changes in the synthesis/levels of the above mentioned substances have been mainly observed during osmotic challenges (12, 13). CRH immunoreactive cells have been demonstrated in the magnocellular parts of the PVN and SON in dehydrated but not euhydrated animals (16). Long term salt loading has been shown to increase CRH- and TH-immunoreactivities in both PVN and SON magnocellular neurons (17) and increased blood plasma osmolality has been shown to stimulate NPY synthesis (18).
Homozygous Brattleboro rats, due to an inability to produce functional AVP (19), suffer from permanent osmotic stress. Therefore di/di rats represent appropriate model of a persisting spontaneous osmotic stimulation which maintains magnocellular neurons in both PVN and SON under constant hypertrophy and hyperactivity (20). In the hypothalamus of di/di rats, basal content of CRH has been found to be similar to that described in Long Evans rats (21). On the other hand, the concentration of NPY has been shown to be 4-fold greater in the neurohypophysis of di/di in comparison with normal Sprague-Dawley rats (22).
In the regulation of PVN and SON magnocellular neurons, a large number of afferent
pathways are involved that utilize a broad range of neurotransmitters (23-26).
The magnocellular neurons receive rich noradrenergic input from the A1
cell group of the lower brainstem (27) and in both PVN and SON, distinct immunoreactivity
for
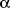 1
1-
and
2-adrenoceptors
has been described (28). Previously, we have shown that: 1) activation of
2-adrenoceptors
by an
2-adrenoceptor
agonist xylazine stimulates approximately 30% of the magnocellular OXY-ergic
neurons in the PVN and SON under normal (control) conditions (29, 30) and 2)
the
2-adrenoceptor
antagonist atipamezole significantly reverses the stimulatory impact of XYL
(31). The goal of the present study was to reveal the involvement of
2-adrenoceptors
in the regulation of magnocellular neurons producing TH, NPY, and CRH under
normal physiological conditions (Long Evans rats) and AVP deficient di/di rats,
permanently suffering from sustained osmotic challenge. To accomplish this goal,
the rats were subordinated to treatments with either
2-adrenoceptors
agonist - XYL, or
2-adrenoceptor
antagonist - ATIP, or both of them. The proper choice of
2-adrenoceptors
agonists and antagonists was made based on the data of our previous studies
(29, 31).
MATERIAL AND METHODS
Animals, treatments, and tissue processing
Adult male Long Evans (n=29) and homozygous Brattleboro rats (n=36) (Harlan, Indianapolis, USA), weighing 250-350 g, were used. The animals were bred in the Institute of Experimental Medicine, Hungarian Academy of Sciences in Budapest (Hungary). Rats were housed one per cage under controlled lighting (light from 7:00 a.m. to 7:00 p.m.) and temperature (22±2°C). Principles of the animal care and all experimental procedures were approved by the Animal Welfare Committee of the Institute of Experimental Medicine which conforms the National Institute of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23) revised 1996.
All the rats were fed with dry food. Control Long Evans rats had free access
to water. To reach more homogenity in the osmotic values, the di/di rats had
regulated water intake during the light period of the day for 4 consecutive
days. They were allowed to drink 15 min at 4 hours intervals,
i.e. at
7:00 and 11:00 a.m. and 3:00 and 7:00 p.m. During the dark period of the day,
the di/di rats had also free access to water. Water intake of control animals
was measured at 7:00 a.m. and 7:00 p.m. and di/di rats every 4 hours during
the light period of the day (7:15 and 11:15 a.m. and 3:15 and 7:15 p.m.) for
four consecutive days. On the day 5
th, different
groups of rats were intraperitoneally (i.p.) injected with: 1/ saline (SAL)
(0.1 ml/100 g BW), 2/ XYL (10 mg/kg), 3/ ATIP (1 mg/kg), and 4/ ATIP followed
5 minutes later by XYL. All the treatments were performed between 7:00 and 11:00
a.m. and after the treatments the animals had no access to water. The selected
time range of treatments (4 hours) was associated with water synchronization
process (which lasted 4 hours), since longer time without water acces might
significantly influence the plasma osmolality values (31). Ninety minutes after
injections, the rats were deeply anesthesized with sodium pentobarbital (50
mg/kg, i.p.) and sacrificed by transcardial perfusion with 50 ml of ice-cold
saline containing 450 µl of heparine (5000 IU/l) followed by 200 ml of fixative
which contained 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4).
The brains were postfixed in the fresh fixative at 4°C overnight and then sunk
in 30% sucrose in 0.1 M PB for 2 days. Thereafter, the brains were frozen in
powdered dry ice and cut into coronal sections of 30 µm thickness on a sledge
microtome (Shandon, Life Sciences International Europe Ltd, UK). The sections
were collected in antifreeze solution and stored at - 20°C until used.
Other important indicators, such as water intake and osmolality of experimental animals, have already been published elsewhere (30).
Single Fos immunohistochemistry
Free floating sections were washed several times in 0.1 M PB and then pre-incubated
with 3% H
2O
2
in 0.1 M PB for 30 min at room temperature (RT). Afterwards the sections were
washed 3 x 10 min in 0.1 M PB and incubated with a polyclonal Fos protein antiserum
(1:1500) diluted in 0.1 M PB containing 4% normal goat serum (Gibco, Grand Island
NY, USA), 0.5% Triton-X 100 (Koch-Light Lab. Ltd., Colnbrook Berks, UK), and
0.1% sodium azide (Sigma, Chemical Ltd., St. Louis, MO, USA) for 48 hours at
4°C. After several rinsings in 0.1 M PB, the sections were incubated with biotinylated
goat anti-rabbit IgG (1:500, VectorStain Elite ABC Kit, Vector Lab., Burlingame,
CA, USA) for 90 min at RT. Next several washings were followed by an incubation
with avidin biotin peroxidase complex (1:250) for 90 min at RT. Fos antigenic
sites were visualized with 0.02% 3,3´- diaminobenzidine tetrahydrochloride (DAB,
Sigma), 2.5% nickel ammonium sulphate (Sigma) and 0.0021% H
2O
2
in 0.05 M sodium acetate buffer (SAB, pH 6.0) for 4-6 min. The heavy metal intensification
of DAB caused black staining of the Fos labeled cell nuclei.
Dual Fos-TH, -NPY, and -CRH immunohistochemistry
After several washings in 0.1 M PB, the Fos immunostained sections were incubated with TH (1:2000), NPY (1:2000), or CRH (1:2000) antibodies for 48 hours at 4°C and afterwards treated by the same way as described above. The sections were stained with single DAB in 0.05 M Tris-HCl buffer (pH 7.4) with 0.06% H2O2 until the TH, NPY, and CRH positive neurons reached yellow - brownish color in the light microsope. Finally, the sections were mounted into 1% gelatine dissolved in 0.0125 M SAB, air-dried, coverslipped with Entellan (Merck), and examined under computerized Leica DMLS light microscope.
Drugs and antisera
Pentobarbital (Spofa, United Pharmaceutical Works, Czech Republic), XYL (Riemser Arzneimittel AG, Germany), and ATIP (gift from the Orion Pharma, Finland) were used. Fos antiserum (No. 94012, provided by Dr. J.D. Mikkelsen, NeuroSearch A/S Ballerup, Denmark) was raised against N-terminal peptide similar to 2-17 of the rat Fos protein according to the protocol described elsewhere (32, 33). TH and NPY antisera were provided by Dr. J.D. Mikkelsen. CRH antiserum GA-13 was a gift from Dr. G. Aguilera (NICHD, NIH, Bethesda, USA).
Data evaluation
Counting of the single TH, NPY, and CRH immunoreactive and Fos-TH, -NPY, and -CRH double labeled neurons within the SON (from Bregma -1.3 to - 1.4) and PVN (from Bregma -1.6 to -1.88) was performed separately on the right and left sides of the PVN and SON on the 30 µm thick serial coronal sections (n=6 sections per animal) employing a computerized light microscope Leica DMLS equipped with a Canon Digital camera (PowerShot S 40). Rat stereotaxic atlas of Paxinos and Watson (34) was used for the determination of anatomical coordinates. The percentage of the activated TH, NPY, and CRH neurons was calculated from the formula: 100 x (number of the TH, NPY, and CRH perikarya displaying Fos signal/total amount of TH, NPY, and CRH immunolabeled perikarya).
Statistics
The number and percentage of activated SON and PVN neurons producing TH, NPY,
or CRH are expressed unilaterally per animal (n=5-6 per group) and section (n=6
per animal). The results are reported as mean±S.E.M. For the statistical evaluation
of the number of neurons and neuronal activities one-way ANOVA (factor strain)
and two-way ANOVA (factors strain and treatment) were applied.
Post hoc
comparisons were done by Tuckey´s
post hoc test. Differences were considered
significant at p<0.05 or p<0.01. Statistical evaluation was performed by SIGMATAT
software for Windows (StatSoft) software.
RESULTS
Basal conditions
Under basal conditions, in the SON, one way ANOVA found significant effect of
the rat strain on the Fos occurrence (F
1,4=28.14,
p<0.05), on the number and activity of TH (F
1,6=8.07,
p<0.01) and NPY (F
1,6=8.87, p<0.01) cells, and
on the activity of CRH neurons (F
1,4=137.17,
p<0.001). Di/di rats had significantly elevated levels of all the proteins detected
(Tuckey´s
post hoc test, p<0.05). In the SON of di/di rats, a significant
increase in the amount of double stained Fos/TH, Fos/NPY, and Fos/CRH neurons
was detected (Tuckey´s
post hoc test, p<0.05).
In the magnocellular PVN, the same phenomenon as in the SON concerning the Fos
occurence (F
1,5=41.11, p<0.05), was observed
and the number of TH and CRH neurons was nearly the same in both rat strains
(TH - 15.3±3.1 +/+ and 16.3±1.8 di/di; CRH 2.0±1 +/+ and 2.9±1.7 di/di). However,
no NPY immunopositive perikarya were found in any of the rat strains studied.
In the PVN, di/di rats had significantly elevated only the activity of TH neurons
compared to +/+ animals (Tuckey´s
post hoc test, p<0.05).
Effect of xylazine
After XYL treatment, two way ANOVA revealed significant influence of the rat
strain on the Fos occurrence in the SON (F
1,10=54.71,
p<0.001) and PVN (F
1,9=84.91, p<0.001). The
number of Fos profiles was higher in the SON and PVN (
Fig. 1) of di/di
rats (SON - 186.9±15.4; PVN - 192.6±13.3) than in +/+ ones (SON - 46.7±0.5;
PVN - 36.8±14.5). XYL treatment did not significantly affect the Fos presence
in the SON and PVN compared to saline controls (
Fig. 1).
 |
Fig. 1. Fos occurrence in
the PVN and SON of +/+ and di/di control (saline treated, SAL) rats. In
the +/+ rats PVN area (A) is demarcated by vasopressin immunoreactive
perikarya with presence of only a small number of Fos profiles. SON area
(C) indicating not clear Fos presence. In the di/di rats PVN (B) and SON
(D) areas are fulfilled with a large number of immunoreactive Fos profiles
as the consequence of the permanent hyperosmotic status of the body fluids. |
In the SON, two way ANOVA revealed effect of the strain (F
1,16=140.2,
p<0.001) on TH neurons activity (
Fig. 2). In the PVN, two way ANOVA found
significant influence on the strain (F
1,14=5.91,
p<0.001) and also treatment (F
1,14=5.56, p<0.05)
on the activity of TH neurons (
Fig. 2). XYL treatment did not significantly
influence the activity of TH neurons in any of the nuclei studied in any of
the rat strain (
Fig. 2).
 |
Fig. 2. Response, expressed
in percentage, of Fos containing TH immunolabeled magnocellular SON and
PVN neurons to treatments with SAL (0.1 ml/100g/bw), XYL (10 mg/kg/bw),
ATIP (1 mg/kg/bw), and ATIP 5 min later followed by XYL. |
In the SON, two way ANOVA found effect of the rat strain (F
1,14=59.41,
p<0.001) on the activity of NPY neurons. However, XYL administration did not
significantly influence the activity of NPY neurons in the SON of +/+ or di/di
rats (
Fig. 3).
 |
Fig. 3. Response, expressed
in percentage, of Fos containing NPY immunolabeled magnocellular SON neurons
to treatments with SAL (0.1 ml/100g/bw), XYL (10 mg/kg/bw), ATIP (1 mg/kg/bw),
and ATIP 5 min later followed by XYL. |
Two way ANOVA revealed effect of the rat strain (F
1,9=148.11,
p<0.001) on the activity of CRH neurons (F
1,9=148.11,
p<0.001) only in the SON (
Fig. 4). The
2-adrenoceptor
stimulation by XYL did not significantly affect the activity of CRH neurons
in +/+ and di/di rats either in the SON or PVN (
Fig. 4).
 |
Fig. 4. Response, expressed
in percentage, of Fos containing CRH immunolabeled magnocellular SON and
PVN neurons to treatments with SAL (0.1 ml/100g/bw), XYL (10 mg/kg/bw),
ATIP (1 mg/kg/bw), and ATIP 5 min later followed by XYL. |
Effect of atipamezole
After ATIP injection, two way ANOVA revealed significant effect of the rat strain
on Fos presence in the SON and PVN (F
1,15=138.33
p<0.001; F
1,15=148.32, p<0.001). In both hypothalamic
nuclei of di/di rats, we observed significantly elevated Fos occurence (SON
- 197.9±3.9; PVN - 184.9±15) in comparison with +/+ rats (SON - 25.7±9.9; PVN
- 17.5±13.7). ATIP did not significantly affect Fos presence in SON (
Fig.
5) and PVN (
Fig. 6) in comparison with SAL injected animals in any
rat strain.
After ATIP administration, two way ANOVA found significant effect of the rat
strain on the activity of TH neurons in the SON (F
1,30=1402.1,
p<0.05) and PVN (F
1,32=22.68, p<0.05). Tuckey´s
post hoc test did not find statistically significant changes in the PVN
or SON of +/+ or di/di rats (
Fig. 2).
After injection of
2-adrenoceptors
antagonist ATIP, two way ANOVA found statistically significant effect of the
rat strain also on the activity of NPY neurons in the SON (F
1,25=59.41,
p<0.05). All groups of di/di rats had significantly higher number of activated
NPY neurons than +/+ animals. ATIP did not significantly change Fos occurrence
in NPY cells in the SON in any strain of the rats studied (
Fig. 3).
In the SON, but not in the PVN, two way ANOVA revealed statistically significant
influence of the strain (F
1,15=33.88, p<0.05)
on the activity of CRH neurons after ATIP application. ATIP did not significantly
influence the activity of CRH neurons either in the SON or PVN of +/+ or di/di
rats (
Fig. 4).
Effect of atipamezole and xylazine
Two way ANOVA detected effect of the rat strain of Fos occurrence in the SON
(F
1,22=232.06, p<0.001) and PVN (F
1,21=146.09,
<0.001). +/+ rats had significantly decreased number of Fos profiles in the
nuclei of SON magnocellular neurons (0.13±0.1) and PVN ones (13±3.7) compared
to di/di rats (SON - 174.8±11.4; PVN - 166.3±24.2; p<0.05). In the SON of +/+
rats, ATIP suppressed the effect of XYL (XYL - 46.7±0.5, ATIP/XYL - 0.13±0.1),
however, Tuckey´s
post hoc test did not find this decrease statistically
significant. ATIP treatment had no significant effect on Fos occurrence in SON
(
Fig. 5) and PVN (
Fig. 6) of XYL injected di/di rats.
 |
Fig. 5. Fos distribution in the SON after SAL, XYL, ATIP, and ATIP-XYL treatments in di/di rats. The quantity of immunoreactive Fos profiles did not reveal any statistically significant differences between the individual groups of the rats. |
 |
Fig. 6. Fos distribution in
the PVN after SAL, XYL, ATIP, and ATIP-XYL treatments in di/di rats. The
quantity of immunoreactive Fos profiles did not reveal any statistically
significant differences between the individual groups of the rats. |
Two way ANOVA found effect of the rat strain on the activity of TH neurons in
the SON (F
1,30=1402.1, p<0.05) and PVN (F
1,32=22.68,
p<0.05). (
Fig. 2).
Two way ANOVA revealed effect of the rat strain on the activity of NPY neurons
in the SON (F
1,25=59.41, p< 0.05). ATIP did
not alter the effect of XYL on the activity of NPY neurons in the SON. Tuckey´s
post hoc test found significantly lower number of activated NPY neurons
in the SON of +/+ rats treated with SAL, XYL, or ATIP than in all groups of
the di/di rats and in +/+ ATIP/XYL injected ones than in XYL and ATIP treated
di/di rats (
Fig. 3).
In the SON, two way ANOVA found significant effect of rat strain (F
1,15=33.88,
p<0.05) on the activity of CRH neurons. Tuckey´s
post hoc test found
significant increase in the activation of CRH neurons in the SON of ATIP treated
di/di rats in comparison with ATIP and ATIP/XYL injected +/+ rats. In the PVN,
no effect of ATIP on XYL treatment was observed (
Fig. 4).
DISCUSSION
The aim of the present study was to examine the involvement of
2-adrenoceptors in the regulation of magnocellular neurons producing NPY, CRH, and TH under normal physiological conditions (Long Evans rats) and permanent spontaneous osmotic challenge (AVP deficient Brattleboro rats). To accomplish this goal, the animals were treated with either
2-adrenoceptors agonist - xylazine, or alpha
2-adrenoceptor antagonist - atipamezol, or both of them.
There is no doubt that in magnocellular AVP and OXY neurons, which are located in the SON and PVN, are involved in the control of body fluid homeostasis. AVP and OXY neurons also co-produce several other peptides and non-peptide putative neurotransmitters, including NPY, CRH, and TH (7, 35), which functional significance is still unknown. The PVN and SON neurons are known to be innervated from hindbrain
1,
2, and A6 catecholaminergic areas (36) and number of studies have documented that norepinephrine (NE) is involved in the regulation of their neurons (37, 38). Previously we have shown (29) that xylazine, a clonidine analoque acting on presynaptic and postsynaptic receptors of the central and peripheral nervous systems as an
2-adrenergic receptor agonist (39), already under the normal (control) conditions stimulates a significant number (around 30%) of OXY neurons in both the PVN and SON (29) what indicates for the importance of
2-adrenergic receptors in the regulation of magnocellular OXY neurons under basal conditions.
TH is co-produced in magnocellular neurons of the PVN and SON together with AVP (10). In di/di rats, we counted more TH immunopositive neurons than in +/+ rats and almost 90% of them was activated. This observation is in concordance with the data of Meister
et al. (40). In the PVN of +/+ and di/di rats we counted approximately the same number of TH immunopositive cells what contradicts with some other observations (41). However, the activity of these neurons in +/+ rats was lower in comparison with di/di rats. Increased activity of TH neurons in hyperosmotic di/di rats corresponds well with the findings of Abramova and co-workers (11) who also observed, paralelly with osmotic rising time, a progressive increase in the number of TH immunoreactive neurons in the SON. Thus there is no doubt that persisting osmotic stress will be the cause of the permanent stimulation of TH magnocellular neurons.
In the present study, XYL did not significantly influence the activity of TH neurons either in the SON or PVN of +/+ or di/di rats. In the PVN of both strain of rats, XYL did not significantly upregulate the activity of TH cells. On the other hand, Abramova and co-workers (42) have found increased TH expression in osmotically stimulated AVP neurons after administration of
1-adrenoceptor antagonist - prazosin, which clearly emphasize the importance of the central catecholaminergic system in the maintanance of increased TH production in AVP neurons. XYL is an agonist of
2-adrenoceptors which are mainly presynaptic and inhibitive and their activation leads to inhibition of the NE release (43). These data may also indicate that
2-adrenoceptors activation should have a comparable effect with
1-adrenoceptors inhibition. We found partial increase of the TH cells only in the PVN. This could mean that XYL, besides
2-adrenoceptors, may activate also
1-adrenoceptors (44), which stimulation has probably opposite effect as their inhibition by prazosin. In the SON of +/+ or di/di rats, XYL did not influence the activity of TH cells. This situation in di/di rats could be explainable by the fact that TH cells had very high activity even under basal conditions, while +/+ ones exhibited little or none TH immunopositive neurons. After administration of
2-adrenoceptors antagonist ATIP, we observed only non-significant partial elevation in the TH cells activity in the PVN in both rat strains and in the SON of +/+ rats. Brady
et al. (45) have demonstrated increased TH mRNA level in the locus coeruleus after long-term administration of
2-adrenoceptors antagonist, idazoxan. However, more specific antagonist ATIP, used in this study, did not significantly alter the activity of TH neurons. These data indicate that
2-adrenoceptors do not probably represent the main regulatory receptor network involved in the regulation of the magnocellular TH neurons.
Another neuropeptide involved in water-salt homeostasis, co-localized with AVP and OXY, is NPY. Exposure of animals to osmotic stimuli (46) have significantly increased NPY mRNA expression and NPY peptide levels in both the SON and PVN. In the present study, an increased number of NPY immunopositive neurons was observed only in the SON of di/di rats and in accordance with the data of Kagotani and co-workers (47), we detected only NPY immunopositive fibers and not immunoreactive perikarya in the PVN. Discrepancy between SON and PVN might probably be caused by higher sensitivity of the SON neurons to osmotic stress (48). We did not find any significant changes in NPY neuronal activity after stimulation or inhibition of
2-adrenoceptors either in the SON or PVN. However, ATIP in +/+ rats partially elevated the activity of NPY cells. In +/+ rats, the effect of ATIP was supressed by XYL what may suggest a role for
2-adrenoceptors in the regulation of activity of NPY neurons.
CRH is known as a key mediator of responses to stress. However, during osmotic stimulation, CRH produced by magnocellular, but not parvocellular neurons (49), can regulate the activity of AVP and OXY neurons (50). Under basal conditions we did not detect any CRH immunopositive neurons in the SON of +/+ rats. As well, Imaki
et al. (14) did not observe any CRH mRNA under basal conditions. However in di/di rats, we detected CRH immunoreactive neurons even under basal conditions and as much as 80% of them were activated. We found increased activity of CRH magnocellular neurons also in the PVN of di/di rats. Our results are in concordance with those of Dohanics
et al. (51), who after long-term osmotic stress comparable with that occurring in AVP deficient di/di rats, detected elevated CRH mRNA levels. Previous studies (52, 53) have demonstrated that NE stimulates parvocellular population of CRH neurons and CRH mRNA expression (54). But it is known, that parvo- and magno-cellular CRH is activated by different stimuli and therefore it can be assumed that catecholaminergic system differently influences the activity of parvo- and magno-cellular CRH neurons. Itoi and co-workers (38), after NE injection into the PVN of rats, did find an increase in the CRH heteronuclear RNA only in its parvocellular subdivision. We did not observe significant changes in CRH neuronal activity after
2-adrenoceptors stimulation or inhibition. In the PVN of +/+ rats, XYL only partially increased the activity of CRH cells but in di/di rats, in both SON and PVN, XYL non-significantly decreased the activity of CRH neurons. As far as XYL may act also
via 1-adrenoceptors, it is possible that they will play also a role in the regulation of the CRH cells and thus it is very likely that different regulation in +/+ and di/di rats can be caused by activation of different receptor systems under basal conditions (+/+ rats) and under permanent osmotic stress (di/di rats).
Based on the present data, we indicate that in contrast to the normal physiological conditions, where the
2-adrenoceptors are involved in the regulation of OXY neurons, they play only a partial role in the regulation of magnocellular neurons producing TH, NPY, and CRH in both the PVN and SON under normal as well as hyperosmotic conditions induced by AVP deficiency (di/di animals).
Acknowledgement:
This research was supported by Vega 2/7003/27 and CE SAS CENDO grants.
Conflict of interests: None declared.
REFERENCES
- Dellmann HD, Rodriguez EM, Pena P, Siegmund I. Immunohistochemical investigation of the magnocellular peptidergic hypothalamo-neurohypophysial system of the rat chronically stimulated by long-term administration of hypertonic saline. Neuroendocrinology 1988; 47: 335-342.
- Klenerova V, Krejci I, Sida P, Hlinak Z, Hynie S. Modulary effects of oxytocin and carbetocin on stress-induced changes in rat behavior in the open-field. J Physiol Pharmacol 2009; 60(2): 57-62.
- Brunton PJ, Arunachalam S, Russel JA. Control of neurohypophysial hormone secretion, blood osmolality and volume in pregnancy. J Physiol Pharmacol 2008; 59: 27-45.
- Kozniewska E, Romaniuk K. Vasopressin in vascular regulation and water homeostasis in the brain. J Physiol Pharmacol 2008; 59: 109-116.
- Wsol A, Cudnoch-Jedrzejewska A, Szczepanska-Sadowska E, Kowalewski S, Puchalska L. Oxytocin in the cardiovascular responses to stress. J Physiol Pharmacol 2008; 59: 123-127.
- Rokaeus A, Young WS, Mezey E. Galanin coexists with vasopressin in the normal rat hypothalamus and galanin's synthesis is increased in the Brattleboro (diabetes insipidus) rat. Neurosci Lett 1988; 90: 45-50.
- Bondy CA, Whitnall MH, Brady LS, Gainer H. Coexisting peptides in hypothalamic neuroendocrine systems: some functional implications. Cell Mol Neurobiol 1989; 9: 427-446.
- Mikkelsen JD, Schmidt P, Sheikh SP, Larsen PJ. Non-vasopressinergic, non-oxytocinergic neuropeptides in the rat hypothalamo-neurohypophyseal tract: experimental immunohistochemical studies. Prog Brain Res 1992; 191: 367-371.
- Skutella T, Weber T, Jirikowski GF. Coexistence of oxytocin and tyrosine hydroxylase in the rat hypothalamus, an immunocytochemical study. J Neural Transm Gen Sect 1993; 94: 55-61.
- Abramova M, Calas A, Thibault J, Ugrumov M. Tyrosine hydroxylase in vasopressinergic axons of the pituitary posterior lobe of rats under salt-loading as a manifestation of neurochemical plasticity. Neural Plast 2000; 7: 179-191.
- Abramova M, Marsais F, Calas A, Thibault J, Ugrumov M. Dynamical study of tyrosine hydroxylase expression and its correlation with vasopressin turnover in the magnocellular neurons of the supraoptico-posthypophysial system under long-term salt loading of adult rats. Brain Res 2002; 925: 67-75.
- Pirnik Z, Mravec B, Kiss A. Fos protein expression in mouse hypothalamic paraventricular (PVN) and supraoptic (SON) nuclei upon osmotic stimulus: colocalization with vasopressin, oxytocin, and tyrosine hydroxylase. Neurochem Int 2004; 45: 597-607.
- Pirnik Z, Kiss A. Fos expression variances in mouse hypothalamus upon physical and osmotic stimuli: co-staining with vasopressin, oxytocin, and tyrosine hydroxylase. Brain Res Bull 2005; 65: 423-431.
- Imaki T, Katsumata H, Miyata M, Naruse M, Imaki J, Minami S. Expression of corticotropin releasing factor (CRF), urocortine and CRF type 1 receptors in hypotalamic-hypophyseal systems under osmotic stimulation. J Neuroendocrinol 2001; 13: 328- 338.
- Larsen PJ, Sheikh SP, Mikkelsen JD. Osmotic regulation of neuropeptide Y and its binding sites in the magnocellular hypothalamo-neurohypophysial pathway. Brain Res 1992; 573: 181-189.
- Kay-Nishiyama C, Watts AG. Dehydration modifies somal CRH immunoreactivity in the rat hypothalamus: an immunocytochemical study in the absence of colchicine. Brain Res 1999; 822: 251-255.
- Dohanics J, Kovacs KJ, Makara GB. Oxytocinergic neurons in rat hypothalamus. Dexamethasone - reversible increase in their corticotropin-releasing factor-41-like immunoreactivity in the response to osmotic stimulation. Neuroendocrinology 1990; 51: 515-522.
- Larsen PJ, Jukes KE, Chowdrey HS, Lightman SL, Jessop DS. Neuropeptide-Y potentiates the secretion of vasopressin from the neurointermediate lobe of the rat pituitary gland. Endocrinology 1994; 134: 1635-1639.
- Valtin H, Schroeder HA, Bernischke K, Sokol HW. Familial hypothalamic diabetes insipidus in rats. Nature 1962; 196: 109-110.
- Ma D, Morris JF. Protein synthetic machinery in the dendrites of the magnocellular neurosecretory neurons of wild-type Long-Evans and homozygous Brattleboro rats. J Chem Neuroanat 2002; 23: 171-186.
- Kjaer A, Knigge U, Bach FW, Warberg J. Impaired histamine- and stress-induced secretion of ACTH and beta-endorphin in vasopressin-deficient Brattleboro rats. Neuroendocrinology 1993; 57: 1035-1041.
- Hooi SC, Richardson GS, McDonald JK, Allen JM, Martin JB, Koening JI. Neuropeptide Y (NPY) and vasopressin (AVP) in the hypothalamo-neurohypophysial axis of salt-loaded or Brattleboro rats. Brain Res 1989; 486: 214-220.
- Vandesande F, Dierickx K. Immuno-cytochemical demonstration of the inability of the homozygous Brattleboro rat to synthesize vasopressin and vasopressin-associated neurophysin. Cell Tissue Res 1976; 165: 307-316.
- Sladek CD, Kapoor JR. Neurotransmitter/neuropeptide interactions in the regulation of neurohypophyseal hormone release. Exp Neurol 2001; 171: 200-209.
- Cole RL, Sawchenko PE. Neurotransmitter regulation of cellular activation and neuropeptide gene expression in the paraventricular nucleus of the hypothalamus. J Neurosci 2002; 22: 959-969.
- Ciosek J, Izdebska K. Thyrotropin-releasing hormone modulates vasopressin and oxytocin synthesis and release from the hypothalamo-neurohypophysial system of different age male rats. J Physiol Pharmacol 2009; 60(2): 63-70.
- Cunningham ET Jr, Sawchenko PE. Anatomical specificity of noradrenergic inputs to the paraventricular and supraoptic nuclei of the rat hypothalamuss. J Comp Neurol 1988; 274: 60-76.
- Aoki C, Go CG, Venkatesan C, Kurose H. Perikaryal and synaptic localization of alpha 2A-adrenergic receptor-like immunoreactivity. Brain Res 1994; 650: 181-204.
- Pirnik Z, Jezova D, Mikkelsen JD, Kiss A. Xylazine activates oxytocinergic but not vasopressinergic hypothalamic neurons under normal and hyperosmotic conditions in rats. Neurochem Int 2005; 47: 458-465.
- Bundzikova J, Pirnik Z, Mikkelsen JD, Zelena D, Kiss A. Activity variations in the hypothalamic oxytocinergic neurons under stimulation of alpha-2 adrenoceptors in osmotically stressed Brattleboro rats. Ann NY Acad Sci 2008; 1148: 154-160.
- Bundzikova J, Pirnik Z, Zelena D, Mikkelsen JD, Kiss A. 2-adrenergic stimulation of oxytocinergic neurons in the hypothalamic magnocellular neurons is influenced by its antagonists in Brattleboro rats. Cell Mol Neurobiol 2009; 29: 1015-1023.
- Woldbye DP, Greisen MH, Bolwig TG, Larsen PJ, Mikkelsen JD. Prolonged induction of c-fos in neuropeptide Y - and somatostatin-immunoreactive neurons of the rat dentate gyrus after electroconvulsive stimulation. Brain Res 1996; 720: 111-119.
- Mikkelsen JD, Vrang N, Mrosovsky N. Expression of Fos in the circadian system following nonphotic stimulation. Brain Res Bull 1998; 47: 367-376.
- Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. New York, Academic Press, 1998.
- Kiss A, Mikkelsen JD. Oxytocin - anatomy and functional assignments: a minireview. Endocr Regul 2005; 39: 97-105.
- Sawchenko PE, Swanson LW. Central noradrenergic pathways for the integration of hypothalamic neuroendocrine and autonomic responses. Science 1981; 214: 685-687.
- Cunningham ET Jr, Sawchenko PE. Anatomical specificity of noradrenergic inputs to the paraventricular and supraoptic nuclei of the rat hypothalamuss. J Comp Neurol 1988; 274: 60-76.
- Itoi K, Helmreich DL, Lopez-Figueroa MO, Watson SJ. Differential regulation of corticotropin-releasing hormone and vasopressin gene transcription in the hypothalamus by norepinephrine. J Neurosci 1999; 19: 5464-5472.
- Perez R, Cox JF, Arrue R. Probable post-synaptic 2 adrenergic mediated effect of xylazine on goat uterine motility. J Vet Pharmacol Ther 1994; 17: 59-63.
- Meister B, Cortes R, Villar MJ, Schalling M, Hokfelt T. Peptides and transmitter enzymes in hypothalamic magnocellular neurons after administration of hyperosmotic stimuli: comparison between messenger RNA and peptide/protein levels. Cell Tissue Res 1990; 260: 279-297.
- Kiss JZ, Mezey E. Tyrosine hydroxylase in magnocellular neurosecretory neurons. Response to physiological manipulations. Neuroendocrinology 1986; 43: 519-525.
- Abramova MA, Ugriumov MV, Calas A. Noradrenergic afferents modulation of tyrosine hydroxylase expression in rat supraoptic nucleus vasopressinergic neuron during ontogenesis. Zh Evol Biokhim Fiziol 2006; 42: 140-145.
- Ma D, Rajakumaraswamy N, Maze M. Alph2-adrenoceptor agonists: shedding light on neuroprotection? Br Med Bull 2005; 71: 77-92.
- Sinclair MD. A review of the physiological effects of alph2-agonists related to the clinical use of medetomidine in small animal practice. Can Vet J 2003; 44: 885-897.
- Brady LS, Gold PW, Herkenham M, Lynn AB, Whitfield HJ Jr. The antidepressants fluoxetine, idazoxan and phenelzine alter corticotropin-releasing hormone and tyrosine hydroxylase mRNA levels in rat brain: therapeutic implications. Brain Res 1992; 572: 117-125.
- Larsen PJ, Mikkelsen JD, Jessop DS, Lightman SL, Chowdrey HS. Neuropeptide Y mRNA and immunoreactivity in hypothalamic neuroendocrine neurons: effects of adrenalectomy and chronic osmotic stimulation. J Neurosci 1993; 13: 1138-1147.
- Kagotani Y, Hashimoto T, Tsuruo Y, Kawano H, Daikoku S, Chihara K. Development of the neuronal system containing neuropeptide Y in the rat hypothalamus. Int J Dev Neurosci 1989; 7: 359-374.
- Giovannelli L, Bloom FE. c-Fos protein expression in the rat subfornical organ following osmotic stimulation. Neurosci Lett 1992; 139: 1-6.
- Pirnik Z, Petrak J, Bundzikova J, Mravec B, Kvetnansky R, Kiss A. Response of hypothalamic oxytocinergic neurons to immobilization stress is not dependent on the presence of corticotrophin releasing hormone (CRH): a CRH knock-out mouse study. J Physiol Pharmacol 2009; 60: 77-82.
- Arima H, Aguilera G. Vasopressin and oxytocin neurones of hypothalamic supraoptic and paraventricular nuclei co-express mRNA for Type-1 and Type-2 corticotropin-releasing hormone receptors. J Neuroendocrinol 2000; 12: 833-842.
- Dohanics J, Kovacs KJ, Makara GB. Oxytocinergic neurons in rat hypothalamus. Dexamethasone-reversible increase in their corticotropin-releasing factor-41-like immunoreactivity in response to osmotic stimulation. Neuroendocrinology 1990; 51: 515-522.
- Kiss A, Aguilera G. Participation of 1-adrenergic receptors in the secretion of hypothalamic corticotropin-releasing hormone during stress. Neuroendocrinology 1992; 56: 153-160.
- Itoi K, Seasholtz AF, Watson SJ. Cellular and extracellular regulatory mechanisms of hypothalamic corticotropin-releasing hormone neurons. Endocr J 1998; 45: 13-33.
- Itoi K, Suda T, Tozawa F, et al. Microinjection of norepinephrine into the paraventricular nucleus of the hypothalamus stimulates corticotropin-releasing factor gene expression in conscious rats. Endocrinology 1994; 135: 2177-2182.