Lactic Acid Bacteria (LAB) such as
Lactobacillus
acidophilus,
Lactobacillus delbrueckii spp. bulgaricus and
Bifidobacteria are important components of the normal intestinal microflora
in humans and animals. These bacteria act as immunomodulators participating
in generation and maintaining of at least three important activities of mucoid
immune system: a) anti-microbial and pro-inflammatory action mediated by Th1-cytokines
such as IL-12, TNF-
alpha and IFN-
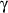
;
b) anti-inflammatory activities and oral tolerance, induced and maintained by
Th2 cytokines, mainly IL-10 and TGF-ß; c) stimulation of adaptive immune
responses, including local and systemic IgG and IgA synthesis being due mainly
to IL-4 and IL-5 (1). Because in humans it would be difficult to create a system
of studying direct interactions between the cells of mucoid immune system and
the comensal bacteria in their natural environment, the use of peripheral blood
mononuclear cells (PBMC) as the responder cells for LAB stimulation
in vitro
is a commonly acceptable alternative. It should be noted that interactions between
commensal bacteria and blood leukocytes may occur due to a limited bacterial
translocation through the epithelial barrier (2, 3). It may also be a reason
for an increase of some non-specific (phagocytosis, Natural Killer cell activities)
and specific (antibody synthesis) immune activities that have been observed
in humans and animals following ingestion of different LAB strains (4). The
cytokines synthesis patterns in the cells of immune system induced by LAB
in
vitro and
in vivo have been shown to be strain-dependent (5, 6).
However little is known how the composition of LAB strains may regulate the
direction and strength of lymphocytes activation reflected either by pro-inflammatory,
antimicrobial defence mechanisms or anti-inflammatory reactivities.
The aim of this study was to find out wheather three LAB strains (
L. acidophilus,
L. bulgaricus and
B. bifidum), acting alone, in pairs or as a mixture would differ from each other in their abilities to induce lymphocytes activities
in vitro measured by lymphocyte proliferation and cytokines synthesis profile.
MATERIAL AND METHODS
Bacteria
The bacteria were prepared according to the procedure described previously (7). We used three bacterial strains: L. delbrueckii ss. bulgaricus LbY-27,
B. bifidum BB 12 R and
L. acidophilus La-5R. The live bacteria strains were used as a stimulating agent in all experiments. The bacterial strains were received from Chr. Hansen, Dairy Ingredients Division, R&D Genetics & Microbiology, Horsholm, Denmark (7).
Isolation of cells
PBMC were obtained from heparinized blood of healthy volunteers (blood donors;
median age 33 years, range 18-55 years), by Isopaque-Ficoll (Lymphoprep; Nycomed
Pharma AS, Oslo, Norway) gradient centrifugation, as previously described (5,
6). Mononuclear cells were resuspended in culture medium made up of RPMI 1640
(Gibco, Paisley, UK) supplemented with 5% heat-inactivated fetal calf serum
(Gibco). Freshly isolated PBMC were diluted in culture medium to a final concentration
of 1×10
6 cells/ml.
Proliferation assay
The assay was made according to the procedure described previously (7, 8). Briefly,
freshly isolated PBMC were diluted in culture medium to a final concentration
of 1×10
6/ml and transfered in a volume of 180
µl to flat-bottomed 96well microtitre trays (Costar, Cambridge, UK). Subsequently,
20 µl of culture medium alone (negative control), 20 µl of Concanavaline A (ConA)
to a final concentration of 50 µg/ml (positive control) or 20 µl LAB in concentration
that induced optimal proliferative response (
L. bulgaricus+
L. acidophilus
- 4×10
6 bacteria/ml, mixture of bacteria and
B.
bifidum 2×10
6 bacteria/ml,
L. bulgaricus+
B.
bifidum,
L. acidophilus+
B. bifidum 1×10
6
bacteria/ml or
L. bulgaricus and
L. acidophilus - 0.5×10
6
of bacteria/ml) were added. The cells were cultured in triplicates for 7 days.
Lymphocyte proliferation was assessed by pulsing the cells with 1 µCi
3H-thymidine
(Amersham, Little Chalfont, UK) for the last 16 h of the incubation period.
The cultures were then harvested onto glass filter strips using an automated
multisample harvester (Skatron, Lier, Norway) and analysed for
3H-thymidine
incorporation by a liquid scintillation counting. Data are given as count per
min (cpm) per 10
6 cells.
Cytokines assay
The assay was performed as previously described (7, 8). Briefly, cytokine concentration
in cell culture supernatants (IFN-
,
TNF-
alpha, IL-12 p40 and IL-10) was determined
after 72 hours of bacterial stimulation of PBMC (1×10
6/ml)
using ELISA (Opt-EIA system, Becton-Dickinson, San Diego, CA, USA), according
to the manufacturers procedure. Dose-response experiments performed for each
cytokine indicated that maximal secretion was obtained with 4×10
6
bacteria/ml (for
L. bulgaricus+
L. acidophilus), 2×10
6
(for mixture of bacteria and
B. bifidum), 1×10
6
(for
L. bulgaricus+
B. bifidum,
L. acidophilus+
B. bifidum)
and 0.5×10
6 (for
L. bulgaricus and
L.
acidophilus), that also induced optimal proliferative response.
Statistics
The STATISTICA (version 6.0) computer software (StatSoft, USA) was used for
the statistical analysis. Values were checked to fulfil the criteria of homogeneity
according to Levene tests. For normal distribution, variables were analysed
by Kolmogorov-Smirnov test with Lillefors correction. For those with non-normal
distribution Mann-Whitney U-test was used. Statistical significance was considered
at
p<0.05.
RESULTS
Proliferative response PBMC
In order to find out if the composition of LAB bacteria strains would have an
effect on their ability to induce lymphocyte proliferation, PBMC were stimulated
with three different LAB strains used together (mixture of bacteria), in combinations
with each other (pairs) or alone, and with a T cell-mitogen Con A, used as a
positive control. All bacterial combinations were equally low inducers of lymphocytes
proliferation (
Fig. 1).
Pattern of cytokine expression in LAB-induced PBMC
The next set of experiments was performed to check if the LAB strains used alone
or in combinations would differ in their capacity for induction of cytokine
pattern in PBMC. Freshly isolated PBMC were activated for 72 hrs with mixture
of three bacterial strains, single LAB strains and their pairs in conditions
as described in
Fig. 1. The amount of cytokines (IFN-
,
IL-12, IL-10 and TNF-
alpha) secreted into the
cell culture supernatants was determined by ELISA.
 |
| Fig. 1.
Proliferative response in PBMC induced by mixture of bacteria, the pairs
of LAB, single LAB strains and Con A (positive control). Values represent
medians obtained from 10 (for mixture of three bacteria strains and L.
acidophilus) and 12 (for remaining composition of LAB) independent
experiments. Mann-Whitney U-test, statistically significant differences:
a - vs. mixture of bacteria, the single and pairs of LAB strains. |
The single LAB strains and their combinations effectively induced different
cytokines synthesis pattern. First,
L. acidophilus was much stronger
inducer of IFN-
and IL-12 than mixture of bacteria,
L. bulgaricus or
B. bifidum.
Second, combination of
L. acidophilus with
L. bulgaricus or
B.
bifidum decreased IFN-
synthesis to the level that was equal to that induced by mixture of bacteria
itself, it also caused a significant reduction of IL-12 production. Third, all
combinations and single strains of LAB were equally good inducers of both IL-10
and TNF-
alpha. The results are presented in
Figs 2-5.
 |
| Fig. 2.
Induction of IFN-
in PBMC stimulated by mixture of bacteria, the pairs or single LAB strains.
Values represent medians obtained from 8 independent experiments. Mann-Whitney
U-test, statistically significant differences: a - vs. L. acidophilus,
b - vs. L. bulgaricus + B. bifidum. |
 |
| Fig. 3.
Induction of IL-12 in PBMC by mixture of bacteria, pairs of LAB strains
and single LAB strains. Values represent medians obtained from 5 (for
single bacteria strains and L. bulgaricus + L. acidophilus)
and 6 (for remaining composition of Lactic Acid Bacteria) independent
experiments. Mann-Whitney U-test, statistically significant differences:
a - vs. mixture of all bacterial strains, b - vs. L. acidophilus,
c - vs. L. bulgaricus, d - vs. L. bulgaricus + L. acidophilus. |
 |
| Fig. 4.
Effect of mixture of bacteria, the pairs and single LAB strains on TNF-alpha
secretion in PBMC. Values represent medians obtained from 8 (for mixture
of all bacteria strains and remaining composition of Lactic Acid Bacteria)
and 5 (for single bacteria strains) independent experiments. Mann-Whitney
U-test, statistically significant differences: a - vs. L. acidophilus,
b - vs. L. bulgaricus. |
 |
| Fig. 5.
Effect of mixture of bacteria, the pairs and single LAB strains on IL-10
secretion in PBMC. Values represent medians obtained from 6 (for all composition
of LAB) and 5 (for single bacteria strains) independent experiments. Mann-Whitney
U-test, statistically significant differences: a - vs. mixture
of bacteria, b - vs. L. bulgaricus. |
DISCUSSION
The study was undertaken to find out if the composition of LAB bacterial strains
used for PBMC stimulation would differ in their effect on: a) the level of lymphocytes
proliferation and, b) the cytokine synthesis pattern. We found that the level
of LAB strains-induced lymphocyte proliferative response in PBMC was equally
low for both the single strains and their combinations (as compared with Con
A-induced controls). The CD4+ T cells were the main responders (not shown) (7).
Concerning this observation, some conflicting results have been reported in
other studies. First, it has been shown that lymphocyte proliferation was increased
in mice fed with different LAB (9) and the T-cell proliferative response
in
vitro to C. albicans,
L. casei GG or B. animalis were stronger in
LAB fed mice than in ones infected with C. albicans alone (10). Second, in humans
consuming fermented milk products supplemented with either
L. acidophilus
La1 or
B. bifidum Bb12, peripheral blood lymphocyte populations have
remained not altered both in term of mitogen-induced proliferation and mode
of surface receptors expression (11). Third,
in vitro studies on human
PBMC stimulated with L. johnsoni La1 and L. sakei LTH have shown moderate proliferative
action of these bacteria in PBMC (12). Altogether these data and our observations
indicate that a moderate proliferative response of lymphocytes to LAB strains
possibly reflect a normal responsiveness of lymphocytes to microflora antigens,
that renders them functionally active but not overstimulated (without harmful,
proinflammatory activities) (
Fig. 1).
In contrast to proliferation, the LAB-induced cytokines synthesis profile in PBMC was strictly dependent on the composition of the LAB strains. First, IFN-
synthesis induced by mixture of bacteria and
L. bulgaricus was equally low but
L. acidophilus induced about 3-fold higher production of IFN-
. Second,
L. bulgaricus and
B. bifidum inhibited
L. acidophilus-induced IFN-
synthesis. Third, the combination of
L. bulgaricus +
B. bifidum resulted in a very low synthesis of IFN-
which was about 4-fold lower than that induced by mixture of bacteria and
L. bulgaricus, and decreased by as much as 10-fold when compared with
L. acidophilus-induced IFN-
secretion (Fig. 2). Down regulation of
L. acidophilus-induced high IFN-
synthesis by the other 2 LAB strains used here may indicate the complex regulation of this cytokine synthesis and confirm anti-inflammatory action exerted by a combination of different LAB strains.
Relatively low mixture of bacteria-induced IFN-
synthesis may reflect the complex mechanisms of control of this cytokine by means of different combinations of LAB that are normally present in the intestine. Rather low activation of IFN-
expression by mixture of LAB strains possibly represents a normal cellular reactivity induced
in vivo by a great variety of LAB strains normally present in digestive tract (13). Their presence is important for maintaining a Th2 profile of immune response which normally prevails in mucosal immune system (14).
This assumption can be further confirmed by the mode of IL-12 expression in
PBMC induced either by mixture of bacteria, the pairs or single LAB strains.
First, mixture of bacteria strains as well as
L. bulgaricus were poor
IL-12 inducers (identically as of IFN-
)
but again,
L. acidophilus strongly induced IL-12 (8fold more than mixture
and 6-fold more than
L. bulgaricus). Second,
B. bifidum was moderate
inducer of IL-12 having about 35% of
L. acidophilus efficacy. Third,
L. bulgaricus and
B. bifidum inhibited
L. acidophilus -mediated
IL-12 synthesis (
Fig. 3).
The data related to the pattern of IFN-
and IL-12 expression in PBMC indicated that particular LAB strains differ in
their capacity for IFN-
and IL-12 induction. It has also been observed by others with respect to other
LAB strains (
L. johnsoni and
L. sakei) which strongly induced
IL-12 and IFN-
(15). These cytokines are known to participate in enhancement of non-specific
immunity and their release may be a reason of increased macrophage phagocytosis
and NK cell activity both in mice injected intraperitoneally with
Lactobacilli
(16), and in humans, following oral administration of
L. acidophilus
or
Bifidobacterium strain Bb12 (17, 18). Since both IL-12 and IFN-
belong to a group of so called proinflammatory cytokines (able to induce harmful
inflammatory reactions), simultaneously participating in immune reactions, its
important to distinguish between inflammation and immunity. Inflammation, which
is the local accumulation of leukocytes, fluid and plasma proteins, is quite
distinct from immunity, defined as the ability to resist infection. So upregulation
of inflammatory mediators (also proinflammatory cytokines) is not necessarily
a desirable attribute for probiotics. Confirmatory to such notion is our finding
that all three of LAB strains studied here showed the ability for strong induction
of IL-10 as well as TNF-
alpha, either used alone
or in combinations (Figs 4-5). The main role of
Lactobacilli-induced
IL-10 would be to limit Th1-mediated proinflammatory response (reflected here
by IFN-
and IL-12 production). In contrast, TNF-
alpha
should participate rather in an enhancement of several cellular defence activities
such as phagocytosis and cytotoxic reactions (20, 21).
On the basis of presented data we conclude that composition of LAB strains is a critical factor which makes possible the enhancement of: 1) cellular activities needed both for down regulation of potential proinflammatory reactions (due to IL-10) and, 2) defence mechanisms necessary for elimination of bacterial and viral pathogens (due to TNF-
alpha expression).
REFERENCES
- Brandtzaeg PE. Current understanding of gastrointestinal immunoregulation and its relation to food allergy. Ann NY Acad Sci 2002; 964: 13-45.
- Deitch EA, Xu DZ, Qi L, Berg RD. Bacterial translocation from the gut impairs systemic immunity. Surgery 1991; 109: 269-276.
- Sedman PC, Macfie J, Sagar P, et al. The prevalence of gut translocation in humans. Gastroenterology 1994; 107: 643-649.
- Wold AE. Immune effects of probiotics. Scand J Nutr 2001; 45: 76-85.
- Miettinen M, Vuopio-Varkila J, Varkila K. Production of human tumour necrosis factor alfa, interleukin-6 and interleukin 10 is induced by lactic acid bacteria. Infect Immun 1996; 64: 5403-5405.
- Maasen CB, van Holten-Neelen C, Balk F, et al. Strain- dependent induction of cytokine profiles in the gut by orally administered Lactobacillus strains. Vaccine 2000; 18: 2613-2623.
- Micha³kiewicz J, Krotkiewski M, Gackowska L, et al. Immunomodulatory effects of lactic acid bacteria on human peripheral blood mononuclear cells. Microb Ecol Health Dis 2003; 15: 185-192.
- Michalkiewicz J, Stachowski J, Barth C, Patzer J, Dzier¿anowska D, Madaliñski K. Effect of Pseudomonas aeruginosa exotoxin A on IFN- synthesis: expression of costimulatory molecules on monocytes and activity of NK cell. Immunol Lett 1999; 69: 359-366.
- De Simone C, Vesely R, Bianchi Salvadori B, Jirillo E. The role of probiotics in modulation of the immune system in man and in animals. Int J Immunother 1993; 9: 23-28.
- Wagner RD, Pierson C, Warner T, et al. Biotherapeutic effects of probiotic bacteria on candidiasis in immunodeficient mice. Infect Immun 1997; 65: 4165-4172.
- Schiffrin EJ, Rochat F, Link-Amster H, Aeschlimann JM. Donnet-Hughes A. Immunomodulation of human blood cells following the ingestion of lactic acid bacteria. J Dairy Sci 1995; 78: 491-497.
- Haller D, Blum S, Bode C, Hammes WP, Schiffrin EJ. Activation of human peripheral blood mononuclear cells by non-pathogenic bacteria in vitro: evidence of NK cells as primary targets. Infect Immun 2000; 68: 752-759.
- Isolauri E, Sutas Y, Kankaanapaa P, Avrilommi H, Salminen S. Probiotics. Effects on immunity. Am J Clin Nutr 2001; 73: 444-450.
- Brandtzaeg P, Baekkevold ES, Farstad IN, et al. Regional specialization in the mucosal immune system: what happens in the microcompartments? Immunol Today 1999; 20: 141-151.
- Miettinen M, Matakainen S, Vuopio-Varkila J, et al. Lactobacilli and Streptococci induce interleukin-12 (IL-12), IL-18, and gamma interferon production in human peripheral blood mononuclear cells. Infect Immun1998; 66: 6058-6062.
- Kato I, Yokokura T, Mutai M. Augmentation of mouse natural killer activity by Lactobacillus casei and its surface antigens. Microbiol Immunol 1984; 28: 209-217.
- Chiang BL, Sheih YH, Wang LH, Liao CK, Gill HS. Enhancing immunity by dietary consumption of a probiotic lactic acid bacterium (Bifidobacterium lactis HN019}: optimization and definition of cellular immune responses. Eur J Clin Nutr 2000; 54: 849-855.
- Sheih YH, Chiang BL, Wang LH, Liao CK, Gill HS. Systemic-immunity enhancing effects in healthy subjects following dietary consumption of the lactic acid bacterium Lactobacillus rhamnosus HN001. J Am Coll Nutr 2001; 20: 149-156.
- Gill HS, Rutherfurd KJ, Cross ML, Gopal PK. Enhancement of immunity in the elderly by dietary supplementation with the probiotic Bidobacterium lactis HN019. Am J Clin Nur 2001; 74: 833-839.
- Strobel S, Mowat AM. Immune response to dietary antigens: oral tolerance. Immunol Today 1998; 19: 173-181.
- Dugas B, Mercenier A, Lenoir-Wijnkoop I, Arnaud C, Dugas N, Postaire E. Immunity and probiotics. Immunol Today 1999; 20: 387-390.