Melatonin influences cell proliferation and differentiation, and the effect of stimulation or suppression of cell division appears to depend on its concentration and cell type examined (10). Anti-proliferative effects of melatonin have been demonstrated in vivo and in vitro in a number of cancer cells (11, 12). In contrast, it stimulates proliferation of normal cells, for example human bone cells (HOB-M) (13) and splenocytes (14).
Melatonin is capable of reducing of free radical damage acting directly as a free radical scavenger, and indirectly, by stimulation of antioxidant enzyme activities (15). It effectively protects against lipid peroxidation and decreases the synthesis of malondialdehyde (MDA) which is an end-product of lipid peroxidation (16, 17). Interestingly, many studies demonstrate that melatonin influences antioxidant enzyme activities under conditions of elevated oxidative stress, for instance after administration free radicals generating agents, such as lipopolysaccharide (LPS), 6-hydroksydopamine or porphyrins (17).
Luzindole (N-acetyl-2-benzyltryptamine) is a relatively selective MT2 receptor antagonist, with a 25-fold higher affinity for the MT2 than for the MT1 receptor subtype. Recent studies revealed that in both in vitro and in vivo conditions, it attenuates the ability of melatonin to enhance splenic lymphocyte proliferation (14, 18), however, there is no information about melatonin effects on other cell types. Thus, the aim of our present study was to examine the influence of melatonin and luzindole on proliferation of preadipocyte 3T3-L1 cells. Moreover, we studied the effect of melatonin on antioxidative enzyme activities and MDA concentration in 3T3-L1 cells media.
Cell culture
3T3-L1 cells are preadipocytes obtained by Green and Kehinde from murine 3T3 fibroblasts by cloning clusters of cells filled with fat droplets (19). 3T3-L1 cells were purchased from ATCC (American Type Culture Collection, Rockville, MD, USA). Preadipocytes were plated at a density of 1 x 106 cells per 25 cm2 flask and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, 100 U/ml penicillin and 0.1 mg/ml streptomycin under an atmosphere of 95% air and 5% CO2 at 37°C.
Experimental protocol
3T3-L1 cells, with or without melatonin, were cultured for 24 or 48 h; and incubation media were not changed during this time. After incubation periods, media were removed, centrifuged and freezed until enzymatic measurements. Cells were trypsinized and number of cells was estimated by direct counting, using a net micrometer with 10x objective and 10x ocular, on three square (1 mm2). Results were calculated as cell number (cells/ml) as average of count per square x 10000. viability of cells was estimated by trypan blue staining. The cell monolayer was discarded, harvested and treated with trypan blue.
Enzymatic assays
Manganese-containing (MnSOD) and copper-zinc-containing superoxide (Cu/ZnSOD) dismutase isoenzyme (EC 1.15.1.1) activities were estimated according to Oyanagui and expressed in nitrite units/ml (NU/ml) (20, 21). Glutathione peroxidase (GSH-Px) (EC 1.11.1.19) activity was measured according to Paglia and Valentine using enzymatic conjunction with glutathione reductase (22). Catalase (CAT) (EC 1.11.1.6) activity was measured according to the kinetic method of Aebi and expressed in IU/ml of medium (23). Glutathione reductase (GSSH-Rd) (EC 1.6.4.2) assay was based on the oxidation of NADPH to NADP+ (24).
MDA concentrations were determined according to the colorimetric method by Ohkawa et al. using the reaction with thiobarbituric acid (25).
Drugs
The following agents were used: melatonin, luzindole - Sigma Chemical CO. (St. Louis, MO, USA), trypsin, trypan blue, Penicillin-Streptomycin Mixture, Dulbecco's Modified Eagle's Medium and Fetal Bovine Serum (Bio Whittaker, Verviers, Belgium). Melatonin was dissolved in a minimum amount of ethanol (95%) and diluted in the medium to obtain a stock solution of 10-3 M/L. Additional dilutions were done in the medium to achieve a final solution of 10-6 M/L. Luzindole was also dissolved in 0.2 ml of ethanol (95%), and then diluted with medium to reach the final concentration of 10-4 M/L. Luzindole was added 30 min before subsequent exposure of the cells to melatonin.
Statistics
All values are given as means ± SEM. Comparison of differences between the mean values were made using ANOVA and Student's t-test. Differences with P < 0.05 were considered as statistically significant.
Influence of melatonin and luzindole on 3T3-L1 cells proliferation
Melatonin at concentrations of 10-3 M/L and 10-6 M/L stimulated proliferation of 3T3-L1 cells in dose- and time-depending manner from 6 to 50% compared to the control (Fig. 1). There were no differences between the control and melatonin-treated groups in regards to viability of the cells.
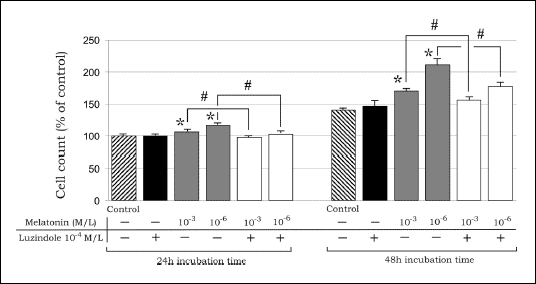 |
| Fig.1. Influence of melatonin (10-3, 10-6 M/L) and luzindole (10-4 M/L) on cell count of 3T3-L1 preadipocytes; data are given as mean ± SEM; * P<0.05 vs. the coresponding value in the control group; in luzindole pretreated groups # P<0.05 vs. melatonin treated groups. |
Luzindole (10-4 M/L) given alone slightly, but not significantly, increased 3T3-L1 cells number in comparison to the control (Fig. 1). In contrast, pre-treatment with this antagonist in the melatonin-treated groups resulted in an inhibition of the pro-proliferative effect of this hormone at 24 and 48 h of incubation (Fig. 1).
Influence of melatonin on antioxidative enzyme activities in 3T3-L1 cells media
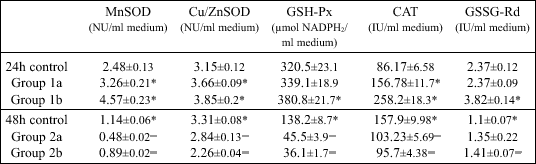
At 24 h of incubation, there were significantly higher media activities of MnSOD, Cu/ZnSOD and CAT detected after treatment with lower melatonin dose, while in media with higher melatonin concentration, the activities of all studied enzymes were significantly increased in comparison to the control (Table I).
| Table I. Activity of manganese-containing (MnSOD) and copper-zinc-containing superoxide dismutase (Cu/ZnSOD), glutathione peroxidase (GSH-Px), catalase (CAT), glutathione reductase (GSSG-Rd) and levels of malondialdehyde (MDA) after 24 and 48h of incubation time with melatonin at concentrations 10-3 M/L (group 1a and 2a, respectively) and 10-6 M/L (group 1b and 2b, respectively) in 3T3-L1 preadipocyte cells culture medium; data are given as mean ± SEM from 6 experiments. |
 |
| *P<0.05 vs.
24h control; |
In contrast, after 48 h of incubation with melatonin (10-3 and 10-6 M/L), activities of all studied enzymes, except for GSSG-Rd activity in melatonin 10-3 M/L concentration medium, were significantly lower than in the corresponding control (Table I).
Influence of melatonin on MDA concentration in 3T3-L1 cells media
There were no differences in MDA concentrations in the studied media after 24 h of incubation, whereas after 48 h of incubation, MDA levels were significantly lower in melatonin-treated cells as compared to the control (Fig. 2). Moreover, the MDA concentrations in the media of control cells after 48 h of incubation were significantly diminished as compared to those determined at 24 h of incubation (Fig. 2).
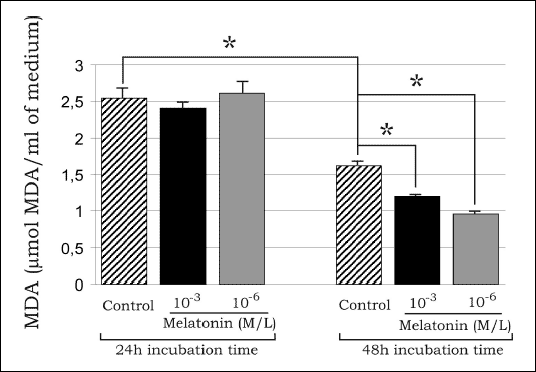 |
| Fig.2. MDA concentrations in media of 3T3-L1 preadipocytes after 24 and 48h of incubation time with melatonin (10-3, 10-6 M/L); data are given as mean ± SEM; * P<0.05 vs. the control group. |
Our results demonstrate for the first time that melatonin acting via MT2 receptors stimulates the proliferation of 3T3-L1 preadipocytes. Moreover, we show increased activities of antioxidative enzymes and decreased lipid peroxidation in melatonin-treated 3T3-L1 cultures.
The cell line employed in our study offers an excellent model to study the differentiation processes. During this process cells undergo a change from the elongated fibroblastic shape to an oval form and accumulate small drops of lipids within the cytoplasm (19). These lipid drops fuse into one large drop giving the cell the aspect of a mature adipocyte of white adipose tissue (19).
Our study demonstrates that melatonin, applied at concentrations of 10-3 and 10-6 M/L stimulates 3T3-L1 cells proliferation in dose- and time-depending manner. Thus, the results confirm earlier findings that melatonin is able to increase the proliferation in normal cells (10). In contrast, melatonin reveals an antiproliferative effect in a number of cancer cells, including those of rat pheochromocytoma PC12 cells (12).
We have shown that MT2 receptor antagonist luzindole (10-4 M/L), added 30 min before exposure of 3T3-L1cells to melatonin, abolished the pro-proliferative effect of melatonin, which suggests the melatonin acting via receptor-dependent mechanism of action. Similarly, melatonin enhanced the proliferative ability of splenocytes in vitro, and this effect was significantly attenuated by luzindole (14, 18). Indeed, our results indicate that also in 3T3-L1 cells melatonin-induced influence on proliferation rate is mediated via MT2 receptors. The particular mechanism involved is not known, especially after results of the study by Sainz et al., who demonstrated that melatonin caused decrease in expression of mRNA for histone H4 in the rat thymus, and in this way, a marked fall in thymocytes proliferation (26).
Results of the present study show that melatonin is able to influence oxidative-antioxidative balance in preadipocytes. Antioxidant properties of melatonin are clearly recognized and they have been studied extensively during recent years (5, 15). Mechanisms of the antioxidant action of this indole include scavenge of radicals and reactive species, induction of antioxidative enzyme activities and inhibition of nitric oxide (NO) synthase activity (17, 27). On the other hand, free radicals are able to alter antioxidant enzyme activities. It has been proposed that moderate levels of toxic oxygen metabolites could induce an increase in antioxidant enzyme activities, while very high levels of these reactants attenuate these enzyme activities as a result of damage of the molecular machinery that is required to induce these enzymes (8). Reactive oxygen species (ROS) are generated in vivo in extremely high amounts during the partial hypoxia and subsequent reoxygenation, for example in haemorrhagic shock (28, 29), or upon exposure to toxic environmental agents, such as those coming from ionizing radiation (30, 31). The experimental evidence confirmed the role of melatonin as a direct ROS scavenger and as an indirect antioxidant, since it stimulates antioxidant enzyme activities (17, 32). Indeed, in the present study, we demonstrated that melatonin affects antioxidative enzyme activities in media after 24 h incubation period of 3T3-L1 cell cultures. A number of studies reported that luzindole abolishes melatonin effects in different species for instance, it produces an augmentation of pancreatic MDA and 4-hydroxynonenal content, and evokes marked increases of plasma levels of lipase, amylase and TNF-alpha in rats with caerulein-induced pancreatitis (33).
Mechanisms of melatonin-induced effects are probably multidirectional. In physiological concentrations (10-9 M/L), it increases mRNA levels of both SOD and GSH-Px isoenzymes in time-dependent manner in rat pheochromocytoma PC12 cells and SK-N-SH neuroblastoma cells (34). Our study is in keeping with this finding by demonstration that melatonin is able to increase activities of MnSOD, Cu/ZnSOD and GSH-Px after 24 h of incubation. Similarly, exogenous melatonin was found to increase the GSH-Px activity in the rat brain, liver and kidney (27).
Under normal conditions, melatonin has no effect on GSSG-Rd activity. However, it protects the cells against a decrease in GSSG-Rd activity produced by okadaic acid (35). Interestingly, in the present study, we have shown that melatonin given at concentration of 10-6 M/L stimulates GSSG-Rd medium activity observed after 24 h of incubation of preadipocytes.
Our results show that melatonin in both concentrations increases CAT activity in 3T3-L1 cell medium after 24 h of incubation. Earlier papers suggested that melatonin produced an enhancement of CAT activity in diabetic skin fibroblasts (36). Moreover, in vivo studies have demonstrated that melatonin caused an increase in CAT activity in rats treated with doxorubicin (37). In addition, it prevents CAT inactivation by neutralization of alkyl-peroxyl radicals (38).
Interestingly, our results demonstrated the fall in activities of all studied enzymes after 48 h of incubation with melatonin, which is difficult to explain. We can only suggest that this effect can be associated with a long-lasting exposure to melatonin and its direct action as a ROS scavenger. Lowered activities of antioxidative enzymes could be related to a decrease in concentration of free radicals generation induced by melatonin.
Free radicals-induced cell damage may be quantitatively determined be measurement of MDA levels, which is an indicator of lipid peroxidation. Present data show that there are no differences in MDA media levels after 24 h of incubation, whereas melatonin had pronounced inhibitory effect on MDA media concentrations after 48 h of incubation. The inhibitory effect of melatonin in MDA levels could by attributed to the increased activities of antioxidative enzymes, which was much more pronounced when 3T3-L1cells were incubated for 48h with this indole.
In conclusion, our results suggest that melatonin stimulates proliferation of 3T3-L1 preadipocytes and that his effect is mediated via MT2 receptors. The enhanced antioxidative enzyme activities and attenuation of the process of lipid peroxidation can contribute to the melatonin-induced increase in proliferation rate observed in 3T3-L1 preadipocytes in vitro.
- Rehfeld JF, Bundgaard JR, Friis-Hansen L, Goetze JP. On the tissue specific processing of procholecystokinin in the brain and gut - a short review. J Physiol Pharmacol 2003; 54: 73-79.
- Peeters TL. Central and peripheral mechanisms by which ghrelin regulates gut motility. J Physiol Pharmacol 2003; 54: 95-103.
- Chey WY, Chang T. Neural control of the release and action of secretin. J Physiol Pharmacol 2003; 54: 105-112.
- Reiter RJ, Tan D, Mayo JC, Sainz RM, Leon J, Bandyopadhyay D. Neurally-mediated and neurally-independent beneficial actions of melatonin in the gastrointestinal tract. J Physiol Pharmacol 2003; 54: 113-125.
- Beyer CE, Steketee JD, Saphier D. Antioxidant properties of melatonin - an emerging mystery. Biochem Pharmacol 1998; 56: 1265-1272.
- Prunet-Marcassus B, Ambid L, Viguerie-Bascands N, Penicaud L, Casteilla L. Evidence for a direct effect of melatonin on mitochondrial genome expression of Siberian hamster brown adipocytes. J Pineal Res 2001, 30: 108-115.
- Bartness TJ, Demas GE, Song CK. Seasonal changes in adiposity: the roles of the photoperiod, melatonin and other hormones, and sympathetic nervous system. Exp Biol Med 2002, 227: 363-376.
- Brydon L, Petit L, Delagrange P, Strosberg AD, Jockers R. Functional expression of MT2 (Mel1b) melatonin receptors in human PAZ6 adipocytes. Endocrinology 2000; 142: 4264-4271.
- Kopecky J, Rossmeisl M, Flachs P, et al. Energy metabolism of adipose tissue - physiological aspects and target in obesity treatment. Physiol Res 2004: S225-232.
- Sainz RM, Mayo JC, Rodriguez C, Tan DX, Lopez-Burillo S, Reiter RJ. Melatonin and cell death: differential actions on apoptosis in normal and cancer cells. Cell Mol Life Sci 2003; 60: 1407-1426.
- Blask DE, Sauer LA, Dauchy RT, Holowachuk EW, Ruhoff MS, Kopff HS. Melatonin inhibition of cancer growth in vivo involves suppression of tumor fatty acid metabolism via melatonin receptor-mediated signal transduction events. Cancer Res 1999; 59: 4693-4701.
- Roth JA, Rosenblatt T, Lis A, Bucelli R. Melatonin-induced suppression of PC12 cell growth is mediated by its Gi coupled transmembrane receptors. Brain Res 2001; 16: 919:139-46.
- Drazen DL, Bilu D, Bilbo SD, Nelson RJ Melatonin enhancement of splenocyte proliferation is attenuated by luzindole, a melatonin receptor antagonist. Am J Physiol Regul Integr Comp Physiol 2001; 280: 1476-1482.
- Nakade O, Koyama H, Ariji H, Yajima A, Kaku T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J Pineal Res 1999; 27: 106-110.
- Reiter JR. Melatonin: lowering the high price of free radicals. News Physiol Sci 2000; 15: 246-250
- Baydas G, Gursu MF, Yilmaz S, et al. Daily rhythm of glutathione peroxidase activity, lipid peroxidation and glutathione levels in tissue of pinealectomized rats. Neurosci Lett 2002; 323: 195-198.
- Rodriguez C, Mayo JC, Sainz RM, et al. Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res 2004; 36:1-9.
- Dubocovich ML. Luzindole (N-0774): a novel melatonin receptor antagonist. J Pharmacol Exp Ther 1988; 246: 902-910.
- Alvarez M. 3T3 cells in adipocytic conversion. Arch Invest Med (Mex) 1991; 22: 235-241.
- Oyanagui Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal Biochem 1984; 142: 290-296.
- Zwirska-Korczala K, Jochem J, Rybus-Kalinowska B, Polaniak R, Birkner E. Assessment of blood superoxide dismutase and glutathione peroxidase activities, and malondialdehyde concentration as oxidation status parameters in obese women. Pol Arch Med Wewn 2003; 110: 725-731.
- Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 1967; 70: 158-169.
- Aebi H. Catalase in vitro. Methods Enzymol 1984; 105: 121-126.
- Carlberg I, Mannervik B. Glutathione reductase. Methods Enzymol. 1985; 113: 484-490.
- Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 1979; 95: 351-358.
- Sainz RM, Mayo JC, Kotler M, Uria H, Antolin I, Rodriguez C. Melatonin decreases mRNA for histone H4 in thymus of young rats. Life Science 1998; 63: 1109-1117.
- Baydas G, Ercel E, Canatan H, Donder E, Akyol A. Effect of melatonin on oxidative status of rat brain, liver and kidney tissues under constant light exposure. Cell Biochem Funct 2001; 19: 37-41.
- Jochem J. Involvement of the renin-angiotensin system in endogenous central histamine-induced reversal of critical haemorrhagic hypotension in rats. J Physiol Pharmacol 2004; 55: 39-55.
- Jochem J, Zwirska-Korczala K, Gwozdz B, Walichiewicz P, Josko J. Cardiac and regional haemodynamic effects of endothelin-1 in rats subjected to critical haemorrhagic hypotension. J Physiol Pharmacol 2003; 54: 383-396.
- Jajte J, Zmyslony M. The role of melatonin in the molecular mechanism of weak, static and extremely low frequency (50 Hz) magnetic fields (ELF). Med Pr 2000; 51: 51-57.
- Ross SM. Combinated DC and ELF magnetic fields can alter cell proliferation. Bioelectromagnetics 1990; 11: 27-36.
- Zwirska-Korczala K, Adamczyk-Sowa M, Polaniak R, et al. Influence of extremely-low-frequency magnetic field on antioxidative melatonin properties in AT478 murine squamous cell carcinoma culture. Biol Trace Elem Res 2004; 102: 227-243.
- Jaworek J, Konturek SJ, Leja-Szpak A, et al. Role of endogenous melatonin and its MT2 receptor in the modulation of caerulein-induced pancreatitis in the rat. J Physiol Pharmacol 2002; 53: 791-804.
- Mayo JC, Sainz RM, Antoli I, Herrera F, Martin V, Rodniguez C. Melatonin regulation of antioxidant gene expression. Cell Mol Life Sci 2002; 59: 1706-1713.
- Tunez I, Munoz MC, Feijoo M, et al. Protective melatonin effect on oxidative stress induced by okadaic acid into rat brain. J Pineal Res 2003; 34: 265-268.
- Kilanczyk E, Bryszewska M. The effect of melatonin on antioxidant enzymes in human diabetic skin fibroblasts. Cell Mol Biol Lett 2003; 8: 333-336.
- Dziegiel P, Murawska-Cialowicz E, Jethon Z, et al. Melatonin stimulates the activity of protective antioxidative enzymes in myocardial cells of rats in the course of doxorubicin intoxication. J Pineal Res 2003; 35: 183-187.
- Mayo JC, Tan DX, Sainz RM, Lopez-Burillo S, Reiter RJ. Oxidative damage to catalase induced by peroxyl radicals: functional protection by melatonin and other antioxidants. Free Radic Res 2003; 37: 543-553.