Cigarette smoke (CS) is presently known as the main cause of cancer, chronic bronchitis, emphysema, cardiovascular disease, and a leading cause of death worldwide (1). CS is also a multipotent carcinogenic mixture that can cause cancer in numerous different organs, and a strong independent risk factor for cancers of oral cavity, upper respiratory tract, lung cancer, and upper gastrointestinal tract (1-3) .
CS contains over 4000 different chemicals, 400 of which are proven carcinogens.
These carcinogens include aromatic amines, nitrosamines, oxidants such as oxygen
free radicals and also high concentrations of toxic volatile aldehydes, such
as acrolein, crotonaldehyde (
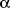
,ß-unsaturated
aldehydes) and acetaldehyde (saturated aldehyde), all of which presumably are
major causes of damage to various molecules exposed to CS (4).
The most abundant aldehyde in CS is acetaldehyde, and its concentration in CS
is >20-80 times greater than that of other aldehydes present in gas phase cigarette
smoke (1). Acetaldehyde is a known mutagen and carcinogen according to various
in vitro cell culture studies and
in vivo animal models. On the
other hand,
,ß-unsaturated
aldehydes such as acrolein and crotonaldyde are even more cytotoxic than are
saturated aldehydes, such as acetaldehyde (5).
Saliva is an exceptional fluid, as it is the first biological fluid to encounter
the inhaled CS gaseous and water soluble extracts. Previous studies have shown
that aldehydes from CS are easily dissolved in the saliva during smoking (6).
Hence, toxic aldehydes could mediate the carcinogenic effect of CS to oral cavity
through saliva and from there, further on to upper respiratory and gastrointestinal
tracts (1). Nagler
et al. (4) have presented evidence that a portion
of the damage to plasma proteins induced by CS was due to its aldehydes contained
in it. In previous studies we have shown that exposure of saliva to plasma
in
vitro resulted in protein modifications, as measured by increased protein
carbonyls (7). The source of the accumulation of protein carbonyls is attributed
mainly to the volatile aldehydes present in CS (4). In addition, it has been
shown that several salivary enzymes such as
-amylase,
lactate dehydrogenase (LDH), and acid phosphatase are considerably affected
by CS, while CS-based aldehydes, such as acrolein and crotonaldehyde, and oxygen
free radicals were implicated as the causative agents affecting those enzymes
(4, 7-9).
Saliva can be considered as one of the chief defense systems against aqueous soluble components of CS (11). As formerly reported, some components of the saliva could neutralize the toxic effect of aldehydes, oxidants, and carcinogens. Saliva contains nonenzymatic and enzymatic antioxidant systems, chiefly superoxide dismutase (SOD), glutathione peroxidase, uric acid, catalase, and glutathione (GSH) (10-13). According to recent studies (4), thiol compounds such as glutathione (GSH) and N-acetylcysteine can provide substantial protection from the CS associated loss of salivary amylase. GSH is possibly involved in the defense mechanism of saliva against noxious CS products, such as aldehydes which react with sulphydryl (-SH) groups. When protein -SH groups are involved, functional impairment of proteins and enzymes will occur. GSH could probably prevent this functional destruction as hypothesized in several
in vitro studies, by making its -SH group available (10).
The purpose of the present study was twofold. Firstly, we sought to determine the interaction of toxic aldehydes with human saliva. Secondly, because thiols have been proposed to have a protective effect against exposure to aldehydes, we examined the protective effects of GSH against aldehyde-induced saliva modifications, and we wished to elucidate the mechanisms whereby CS aldehydes might influence
-amylase activity.
MATERIAL AND METHODS
The study was approved by an institutional Ethics Committee and all volunteers gave their informed consent.
Collection of saliva
Whole saliva, which is basically total oral fluid, was collected from healthy male and female nonsmokers under non-stimulatory conditions in the morning. For the collection, which was performed at least 1 h after eating, volunteers were asked to generate saliva in their mouths and to spit it into a wide test tube for 10-15 min. Following collection, saliva was immediately centrifuged (1000 x g, 3 min) to remove squamous cells and cell debris. The fresh supernatant was immediately used for CS and aldehydes studies, and
-amylase activity studies were performed at the end of the CS and aldehydes exposure studies.
Cigarettes and aldehydes
The cigarettes used in this study were popular commercial cigarettes (Time cigarettes; Dubek Ltd., Tel Aviv, Israel) containing 14 mg of tar and 0.9 mg of nicotine per cigarette. Acrolein was purchased from Sigma Chemicals Co., acetaldehyde from Fisher Scientific Co, and Crotonaldehyde from Aldrich Chemical Co.
Exposure of saliva to CS
An
in vitro study was carried out using Time cigarettes combined with a vacuum system, as described previously (4, 9). Saliva (45 ml) was placed in 250 ml flasks with a sidearm to which the cigarettes were attached. A reproducible vacuum was created in the flask, and after opening the vacuum to the lighted cigarette for 5 s, 80 to 100 ml of CS puffs were drawn into the flask. Flasks were incubated in a metabolic shaker for a total of 3 h at 37°C. In a given experiment, puffs were administered to the flask over a 3-h time period at 20-min intervals, for a total of nine puffs. Samples for biochemical analysis were removed from the flasks at zero time and at 1, 2, and 3 h.
Exposure of saliva to purified aldehydes
4-5 ml of human saliva was treated with an aldehyde mixture of crotonaldehyde and acrolein similar to that reported to be present in a mainstream smoke of a single 2R1 University of Kentucky reference cigarette (5), to give a final concentration of 0.8 µmol/4 ml acrolein and 0.21 µmol/4 ml crotonaldehyde (AL). Another 4-5 ml of saliva was treated with acetaldehyde (ACET-AL) to give a final concentration of 20 µmol/4 ml. Aldehydes were administered to the flask over a 3-h time period at 20-min intervals, for a total of nine times. Samples for biochemical analysis were removed from the flasks at zero time and at 1, 2, and 3 h. Flasks were incubated in a metabolic shaker for a total of 3 h at 37°C.
-amylase activity determination
The AMY assay method using Dimension
® clinical
chemistry system was employed in this project. The AMY method utilizes a chromogenic
substrate, 2-chloro-4-nitrophenol linked with maltotriose. The direct reaction
of
-amylase with
the substrate results in the formation of 2-chloro-4-nitrophenol which is monitored
spectrophotometrically. The AMY method responds to both pancreatic and salivary
amylase isoenzymes. Enzyme activities were expressed as IU/L.
Thermochemiluminescence analysis (TCL)
Photon emission during heating was measured by TCL Photometer (manufactured by Lumitest, Israel) in the saliva, as previously described using a photomultiplier model R265P (Hamamatsu Photonics Co., Japan) with a spectral response range of 300-650 nm. The TCL device measures the level of photon emission from excited carbonyls in biological macromolecules,
i.e., oxidative modifications of lipids and proteins. The fluid examined was distributed over the surface of an aluminum tray (a type of miniature Petri dish) inside the sample preparation block and was then vacuum dried. The dish is then mounted on a constant temperature heater kept at 80° in the analysis block and the photon emission is measured. The TCL curve obtained is described mathematically by means of the two main parameters examined in the analysis (1) the amplitude of the kinetic curve of the photon emission and (2) its slopes. The TCL amplitudes both at 50 s (H1) and 280 s (H3) and the oxygenation potential and preincubation values were recorded, all representing the level of oxidative stress for which the saliva had been exposed to. H1 and H3 are base line and end point parameters of the TCL kinetic curve respectively that reflect electronically excited species formation due to heat induced oxidative modifications (14).
Addition of GSH to saliva
GSH was added to saliva at zero time and flasks were incubated in a metabolic shaker for a total of 30 min at 37° C, prior to the first smoking puff or aldehyde addition, at concentrations of 1mM (GSH from Sigma Chemicals). In addition, in all experiments, a control without the compounds was run with no difference in the assay conditions.
Statistical analysis
Statistical analysis was performed using an unpaired t-test. To determine statistical significance, the ranges, means, and SD were computed. Results are reported as means ±SD. Statistical significance was set at P<0.05.
RESULTS
Fig. 1 shows the loss of
-amylase
activity after exposure of saliva of three volunteers to nine puffs of CS over
a 3-h time period. It can be seen that
-amylase
activity decreased over time by about 30% of the control level (P<0.001). The
initial mean level from the three volunteers was 45x10
3
±8 IU/L.
-amylase
activity did not change when saliva was incubated at 37° C under air. However,
exposure of plasma to increasing numbers of puffs of gas phase CS produced a
steady decrease in
-amylase
activity. We received a similar pattern in response to CS, when a,b-unsaturated
aldehydes, acrolein and crotonaldyde, were added. There was a reduction of 32%
in comparison with the 4.7% reduction in the control (P<0.001). There was no
statistical difference between CS and
,ß-unsaturated
aldehydes in reducing
-amylase
activity. Acetaldehyde also reduced
-amylase
activity in a significant manner in comparison with control (14%
vs.
5%; P<0.001), but in comparison with CS and
,ß-unsaturated
aldehydes it had significantly lower effect in decreasing
-amylase
activity (14%
vs. 33% and 32%; P<0.001 and P<0.02, respectively).
 |
| Fig. 1.
Effects of gas phase cigarette smoke (CS) on amylase activity in saliva.
Salivary amylase activity was measured in the absence of CS or aldehydes
- control (rhomb), in the presence of CS (square), crotonaldehyde & acrolein
(AL; triangle), and acetaldehyde (circle). The results are expressed as
means ±SD percentage of reduction in amylase activity during 3 h and are
the means of eight separate experiments from 3 subjects. *P<0.001; see
statistical explanation in the text. |
Fig. 2 shows changes in saliva TCL ratio. This ratio allows us to observe
oxidative modification of proteins and lipids from time 0 to 3 h. There was
no change in TCL ratio when saliva was incubated at 37° C under air. Exposure
of saliva to CS and
,ß-unsaturated
aldehydes produced a similar increase in TCL ratio (42% ±8 and 44% ±10, respectively).
The increase was significantly different from the 2.0 ±1.5% control level (P<0.001).
Acetaldehyde addition also produced a significant increase in TCL ratio which
was significantly higher than the control (10.0 ±3.3%
vs. 2.0 ±1.5%;
P<0.001), but was lower than the effect of CS and
,ß-unsaturated
aldehydes (P<0.001). As mentioned, CS contains a number of constituents (
e.g.,
oxidants, aldehydes) that can cause protein modifications and influence protein
enzyme activity (15). To elucidate which constituents could cause loss of
-amylase
activity, the saliva was exposed to CS and volatile aldehydes in the presence
of 1 mM GSH.
 |
| Fig. 2.
Salivary TCL. Control on air (square), cigarette smoke (CS; rhomb), crotonaldehydes
& acrolein (AL; circle) and to acetaldehyde (triangle). The results are
expressed as means ±SD percentage of change in the TCL ratio during 3
h and are the means of eight separate experiments from 3 subjects. *P<0.001;
see statistical explanation in the text. |
Fig. 3 shows that the rate of loss of
-amylase
activity was influenced by the presence GSH (33 ±11% for CS
vs. 22 ±7%
for CS+GSH; P<0.05) after 3 h. The addition of the major aldehydes present at
concentrations contained in the CS of one cigarette influenced salivary
-amylase
activity.
Fig. 4 shows that in the setting of added
,ß-unsaturated
aldehydes, the rate of loss of
-amylase
activity was also influenced by the presence of GSH (32 ±10% for
,ß-unsaturated
aldehydes
vs. 14 ±3% for a,b-unsaturated aldehydes+GSH; P<0.001) after
3 h.
Fig. 5 shows that in the setting of added saturated acetaldehydes,
the rate of loss of
-amylase
activity was not significantly influenced by the presence of GSH (14 ±4% for
acetaldehydes
vs. 19 ±8% for +GSH; P<0.001) after 3 h.
 |
| Fig. 3.
Effects of 1mM GSH on salivary amylase activity exposed to cigarette smoke
(CS; rhomb) alone and to CS+GSH (square). The results are expressed as
means ±SD percentage of reduction in amylase activity during 3 h and are
the means of eight separate experiments from 3 subjects. *P<0.05; see
statistical explanation in the text. |
 |
| Fig. 4.
Effects of 1mM GSH on salivary amylase activity exposed to crotonaldehydes
& acrolein (AL; triangle) alone and to AL+GSH (square). The results are
expressed as means ±SD percentage of reduction in amylase activity during
3 h and are the means of eight separate experiments from 3 subjects. *P<0.001;
see statistical explanation in the text. |
 |
| Fig. 5.
Effects of 1mM GSH on salivary amylase activity exposed to acetaldehyde
alone (square) and acetaldehyde+GSH (rhomb). The results are expressed
as means ±SD percentage of reduction in amylase activity during 3 h and
are the means of eight separate experiments from 3 subjects. *P<0.001;
see statistical explanation in the text. |
Fig. 6 is a summary of all the experiments presented in this article. It shows
the similar effects of CS and
,ß-unsaturated
aldehydes on amylase activity and that they both are influenced by the presence
of GSH. It is worth noticing that the saturated aldehyde, acetaldehyde, had
a much smaller effect on
-amylase
activity in comparison with CS and a,b-unsaturated aldehydes (P<0.001; P<0.02)
and that GSH had no influence on acetaldehyde-induced damage to
-amylase
activity, as opposed to GSH profound protective effect against the toxicity
induced by CS and
,ß-unsaturated
aldehydes.
 |
| Fig. 6.
Effects of 1mM GSH on salivary amylase activity exposed to cigarette smoke
(CS) and purified aldehydes. The results are expressed as means ±SD percentage
of reduction in amylase activity after 3 h and are the means of eight
separate experiments from 3 subjects. *P<0.001 **P<0.05; see statistical
explanation in the text. |
DISCUSSION
Many noxious compounds in CS modify biological molecules by acting as oxidants, proinflammatory agents, or carcinogens (16-18). One type of such molecules are unsaturated and saturated aldehydes, present in CS (5, 13). All these toxic compounds first encounter oral cavity tissues, where saliva should act as a protector against the water soluble fraction of CS (10). In the present study, noxious effects of CS aldehydes have been evaluated
in vitro directly in fresh human saliva samples. The major aim of the study was to try to understand the underlying causes behind the phenomenon that gas-phase CS can rapidly inactivate a large percentage of
-amylase activity
in vitro. Firstly, we examined the degree to which volatile aldehydes present in CS could account for the effects of CS on saliva through
-amylase activity and carbonyl formation. Secondly, we examined the ability of GSH to amend the effects of these aldehydes in fresh human saliva.
Previous studies have shown that the aldehydes of CS gas phase could explain
partially some of the protein modification caused by CS. Protein modification
was examined through carbonyl formation and enzymatic activity. Reznick
et
al (7) and ONeill
et al (5) have shown that exposure of plasma proteins
to CS or directly to aldehydes induced their modification as evidenced by the
enhancement of plasma carbonyl groups and a parallel decrease in -SH groups.
In the present saliva study, the effects of CS and volatile aldehydes on protein
carbonyls resemble, to a great extent, those found in plasma studies (7). Exposure
of saliva to a mixture of purified unsaturated aldehydes (crotonaldehyde and
acrolein), at concentrations known to be present in CS, resulted in a similar
increase of protein carbonyls to CS, measured by TCL technology, which measures
excited carbonyls (5). On the other hand, the saturated aldehyde, acetaldehyde,
which is a major volatile constituent of the CS and present in CS in relatively
high concentrations, had a less significant effect on carbonyl formation, as
opposed to CS and unsaturated aldehydes (
Fig. 2).
A more extensive study was performed in order to understand the factors involved
in
-amylase inactivation
due to CS exposure. Exposure of saliva to CS significantly accelerated
-amylase
loss of activity. A similar pattern was shown by the addition of unsaturated
aldehydes, and, to a much less extent, by acetaldehyde (
Fig. 1).
GSH
In previous studies, functional modifications induced by CS were partially prevented
by adding to the incubation system sulphydryl compounds, like GSH, whose role
could be to offer SH groups to aldehydes instead of proteins (10). In other
studies, where purified aldehydes were shown to cause accumulation of protein
carbonyls in plasma (10), GSH was capable of ameliorating this accumulation
in both plasma and salivary proteins (4, 10). In the present study, GSH slowed
down the rate of CS-associated loss of
-amylase
activity (
Fig. 3). A comparable influence was found when saliva was incubated
with GSH before the addition of unsaturated aldehydes (
Fig. 4). There
was no significant influence in the case of acetaldehyde (
Fig. 5). GSH
did not completely inhibit the CS and unsaturated aldehydes decrease of
-amylase
activity. However, GSH slowed this loss considerably. This may suggest that
GSH provides protection against
-amylase
oxidation, while protecting some protein -SH groups, probably at the active
site of the enzyme. Indeed, the role of -SH groups in preserving
-amylase
activity has already been shown (4).
The present study indicates that aldehydes present in CS, especially unsaturated aldehydes, contribute significantly to protein modification caused by CS and that thiols appear to be protective against aldehyde-induced protein modification as measured by the end points of excited carbonyl formation (TCL) and
-amylase activity. It is likely that thiols such as GSH are potentially beneficial in protecting proteins against the toxic effects of unsaturated aldehydes.
Proposed mechanism
The results of the present study suggest that the unsaturated aldehydes in CS are the major source of reactive double bonds reacting with -SH groups of proteins in the Michael addition reaction, attaching the aldehydic carbonyls to the proteins (4, 5). Indeed, in previous studies in plasma, it was shown that a 1:1 stoichiometry, on a molar-to-molar basis, exists between the appearance of protein carbonyls and the disappearance of protein thiol groups (5). Thus, it is quite reasonable to assume that also in the case of saliva, the increase of protein carbonyls, due to exposure to CS, is mainly due to reaction of unsaturated aldehydes with protein -SH groups, adding their carbonyls attached to proteins (4).
The mechanism of protein damage by saturated aldehydes is more bewildering.
It is possible that these aldehydes react with proteins, forming adducts at
both -SH and -NH
2 groups, but it is unlikely
that these adducts will contain carbonyl groups. Therefore, the increase in
carbonyl formation after the addition of acetaldehyde was significantly lower
in comparison with CS and unsaturated aldehydes.
The reduction of aldehyde-induced protein modification by GSH may stem from the added -SH moieties that are available to react with aldehydes. Indeed, the addition of GSH to saliva, may reduce the concentration of mixed disulfides, thus increasing the amount of protein-SH moieties, suggesting that unsaturated aldehyde-induced protein damage could be ameliorated by augmented GSH (5).
The results of our study clarify the underlying mechanisms of the immediate and overwhelming induced inactivation of
-amylase by CS and aldehydes, and of partial protection rendered by GSH against it. Our data show that the addition to saliva of a mixture of aldehydes found in CS leads to the formation of products that are detectable in the carbonyl assay and cause some inactivation of
-amylase. Therefore, we conclude that at least some of the carbonyl formation and
-amylase inactivation caused by CS is due to aldehydes, chiefly unsaturated aldehydes.
The loss of
-amylase activity and carbonyl formation reported in the present study, along with other studies due to CS and aldehydes, could play an important role in the initiation and progression of oral inflammatory diseases, promote premalignant and malignant lesions of the oral mucosa, and destroy normal homeostasis of the oral cavity. We found that treating the whole saliva with antioxidant thiols, such as glutathione, shows that GSH might be a significant protector against CS and/or unsaturated aldehyde-induced damage. Further studies are needed to elucidate the role of CS-related inhibition of saliva
-amylase activities in oral pathobiology, If corroborated by further studies, these findings may help develop an effective means of protecting against CS-associated damage to salivary enzymes and proteins.
Acknowledgment:
This work was supported by the Krol Foundation of Barnegat, NJ, USA and a grant
from the vice president of research of the Technion.
Conflicts of interest: the authors declare no conflicts of interest in
relation to this work.
REFERENCES
- Salaspuro VJ, Hietala JM, Marvola ML, Salaspuro MP. Eliminating Carcinogenic Acetaldehyde By Cysteine From Saliva During Smoking. Cancer Epidemiol Biomarkers Prev 2006; 15: 146-149.
- Anonymous. Tobacco smoking and involuntary smoking. IARC monographs on the evaluation of carcinogenic risk of chemicals to humans. 83. Lyon (France): IARC; 2004.
- Sasco AJ, Secretan MB, Straif K. Tobacco smoking and cancer: a brief review of recent epidemiological evidence. Lung Cancer 2004; 45 Suppl: 53-59.
- Nagler R, Lischinsky S, Diamond E, Drigues N, Klein I, Reznick AZ. Effect of cigarette smoke on salivary proteins and enzyme activities. Arch Biochem Biophys 2000; 379: 229-236.
- ONeill CA, Halliwell B, Van der Vliet A et al. Aldehyde induced protein modification in human plasma: Protection by glutathione and dihydrolipoic acid. J Lab Clin Med 1994; 124: 359-369.
- Salaspuro V, Salaspuro M. Synergistic effect of alcohol drinking and smoking on in vivo acetaldehyde concentration in saliva. Int J Cancer 2004; 111: 480-483.
- Reznick, AZ, Cross CE, Hu ML et al. Modification of plasma proteins by cigarette smoke as measured by protein carbonyl formation. Biochem J 1992; 286: 607-611.
- Klein I, Nagler RM, Toffler R, Van der Vliet A, Reznick AZ. Effect of cigarette smoke on oral peroxidase activity in human saliva : Role of hydrogen cyanide. Free Radic Biol Med 2003; 35: 1448-1452.
- Nagler RM, Lischinsky S, Diamond E, Klein I, Reznick AZ. New insights into salivary lactate dehydrogenase of human subjects. J Lab Clin Med 2001; 137: 363-369.
- Zappacosta B, Persichilli S, Mordente A et al. Inhibition of salivary enzymes by cigarette smoke and the protective role of glutathione. Hum Exp Toxicol 2002; 21: 7-11.
- Mandel ID. The functions of saliva. J Dent Res 1987; 66: 623-627.
- Tabak LA, Levine MJ, Mandel ID, Ellison SA. Role of salivary mucins in the production of oral cavity. J Oral Pathol 1982; 11: 1-17.
- Moore S, Calder KA, Miller NJ, Rice-Evans CA. Antioxidant activity of saliva and periodontal disease. Free Radic Res 1994; 21: 417-425.
- Reznick AZ, Shehadeh N, Shafir Y, Nagler RM. Free radicals related effects and antioxidants in saliva and serum of adolescents with Type 1 diabetes mellitus. Arch Oral Biol 2006; 51: 640-648.
- Bar-Shai M, Hasnis E, Wiener-Megnazi Z, Reznick AZ. The role of reactive nitrogen species and cigarette smoke in activation of transcription factor NF-kB and implication to inflammatory processes. J Physiol Pharmacol 2006; 57 Suppl 4: 39-44.
- Pryor WA, Prier DG, Church DF. Electron spin resonance study of main mainstream and sidestream cigarette smoke: nature of the free radicals in gas-phase smoke and in cigarette tar. Environ Health Perspect 1983; 47: 345-355.
- Hasnis E, Bar-Shai M, Burbea Z, Reznick AZ. Cigarette smoke-induced NF-kB activation in human lymphocytes: the effect of low and high exposure to gas phase of cigarette smoke. J Physiol Pharmacol 2007; 58 Suppl 5: 263-274.
- Hasnis E, Bar-Shai M, Burbea Z, Reznick AZ. Mechanisms underlying cigarette smoke-induced NF-kB activation in human lymphocytes: the role of reactive nitrogen species. J Physiol Pharmacol 2007; 58 Suppl 5: 275-288.