The process of immunoageing, accompanied by a progressive remodeling of the function of the immune system, is to a great extent regulated by different hormonal immunomodulators such as estrogens, androgens, progesterone, glucocorticoids and melatonin which blood levels and cellular transduction systems undergo significant changes in the elderly (1-3). Ageing of the immune system relates to both innate and adaptive immunity although the innate one is regarded to be better preserved than adaptive one. The age-related alterations of adaptive immunity include a decline of naive T lymphocytes and accumulation of memory/effector T cells, reduction in the number of B cells, up-regulation of the inflammatory responses and dysregulation of the Th1/Th2 system. The increased number of NK cells with well-preserved cytotoxic function is a characteristic feature of the innate part of ageing immune system (4).
NK cells (CD3
- CD16
+
CD56
+) are important components of antiviral and
antitumor immunity (5). Morphologically they represent large granular lymphocytes
and constitute approximately 10-20% of peripheral blood lymphocyte population.
The NK activity well correlates with a health status. Healthy individuals may
reveal high cytotoxic activity until late senescence and present cytotoxic capacity
similar to the young adult counterparts (6, 7). Contrary to other cells of innate
immunity, activated NK cells can proliferate and low telomerase activity has
been detected even in non-activated NK cells (8, 9). Mature granulocytes, monocytes
and mast cells do not proliferate and do not reveal telomerase activity (10).
Telomere shortening is one of the hallmarks of the ageing process and results from inability of DNA polymerases to replicate a linear DNA molecule (end replication problem) and presence of unrepaired damage of DNA caused mainly by oxidative stress (11). Telomeres are structures located at the ends of eukaryotic chromosomes composed of tandemly repeated short sequences (TTAGGG in vertebrates) associated with specific proteins (12). They progressively shorten with each cell division that results in most somatic cells in chromosomal instability and cellular senescence (11). Similarly to other cells, telomeres of the immune cells including B and T lymphocytes, granulocytes, monocytes and NK (natural killer) cells shorten with age (9, 10). Activated lymphocytes, however, increase the activity of telomerase, a ribonucleoprotein DNA polymerase complex that maintains telomere length. The complex comprises the protein reverse transcriptase (hTERT) and a template RNA (hTR). Telomerase activity is absent in most normal human somatic cells because of the lack of hTERT expression. Normally telomerase activity is detected only in stem cells, germ line cells and activated lymphocytes (13). Telomerase activity is also found in neoplastic cells (14).
The process of successful ageing,
i.e. ageing in good psychophysical conditions (15) is immunologically characterized by preserved lymphoproliferative responses and NK cytotoxic activity as well as conserved antigen presentation (16). Healthy centenarians show normal number of T lymphocytes, increased production of immunoglobulins, lack of organ-specific autoantibodies, well preserved NK activity and retained proliferative capability of T lymphocytes (17). Studies on human longevity performed by Olshansky
et al. revealed that the age of 85 years may be regarded as the average biological limit to life (18). Studies on the phenomena of longevity and successful ageing performed on groups of seniors at the age over 85 are therefore of special interest.
The aim of our study was to compare the function of NK cells, their cytotoxic properties and sensitivity to activation in the group of nonagenarians representing population of selected, long-lived seniors and a group of elderly below the critical age of 85, who belonged to much larger population of seniors. Moreover, we measured telomere length and telomerase activity in NK cells as parameters of cellular ageing.
MATERIAL AND METHODS
Study population
Peripheral blood mononuclear cells (PBMC) were obtained from 35 young subjects referred to as young (mean age 25±4 ys; 19 women and 16 men) and 35 elderly: 28 seniors at the age below 85 referred to as old (mean age 78±5 ys; 23 women and 5 men) and 7 seniors at the age over 85 referred to as the oldest (mean age 92±2 ys; all women). The elderly volunteers did not suffer from infections, inflammatory or autoimmune diseases for 6 months before and during the study. However, some seniors were afflicted with degenerative joint and spine disease (14), cataract (3), depression (7), circulatory insufficiency (4), ischemic heart disease (10), ischemic brain disease (2), diabetes mellitus (4), osteoporosis (3). Informed consent from volunteers and approval from Ethical Committee of Medical University of Gdansk were obtained.
Peripheral blood mononuclear cell separation
Peripheral blood mononuclear cells (PBMC) were isolated from heparinised peripheral blood by conventional ficoll-uropoline density gradient centrifugation. PBMC were collected at the interphase, washed and resuspended in PBS-1% FBS solution.
NK cell separation
NK cells (CD3
- CD16
+
CD56
+ cells) were isolated from PBMC by negative
selection with the use of NK isolation kit and MACS magnetic separator (Miltenyi
Biotec, USA). PBMC were incubated with Biotin-Antibody Cocktail (suspension
of biotin-conjugated monoclonal antibodies against CD3, CD4, CD14, CD15, CD19,
CD36, CD123 and Glycophorin A) for 10 min, then washed with washing buffer and
incubated with Anti-Biotin Microbeads (microbeads conjugated to monoclonal antibiotin
antibodies) for subsequent 15 min. Then cells were resuspended in 500 µl of
washing buffer and applied onto the column. Cells eluted from the column in
magnetic field represented the enriched NK cell fraction of about 90% purity.
The purity of the enriched NK cells was evaluated by flow cytometry. Aliquots
of the cell fractions were stained with anti-CD56 PE-conjugated (BD, PharMingen,
USA) and anti-CD3 FITC-conjugated (BD PharMingen, USA) monoclonal antibodies
(20 µg/test).
NK cytotoxic activity
PBMC (2x10
5 cells/200 µl) were cultured with target
K562 cells (2x10
4 cells/200 µl) mixed in a ratio
of 10:1 in RPMI 1640 medium supplemented with 1% FBS for 4 h. Then culture plates
were centrifuged and supernatants were transferred to new plates. Subsequently
the amount of lactate dehydrogenase (LDH) released from the lysed cells was
estimated with the use of Cytotoxicity Detection Kit (Roche Diagnostics, Germany).
LDH present in supernatants reduced tetrazolium salt to formazan salt resulting
in a color reaction. The absorbance of samples was measured at 490 nm with microplate
reader (Multiscan MCC/340, Labsystems, Finland). Spontaneous LDH release was
determined by incubation of target cells with culture medium. The maximum release
of LDH was determined by incubation of target cells in the medium with a final
concentration of 1% Triton X-100 (Sigma Chemical, USA). The percentage of NK
cytotoxicity was calculated according to the formula: NK cytotoxicity (%) =
[(experimental value spontaneous LDH release) / (maximum LDH release spontaneous
LDH release)] x 100%.
Detection of NK cells (CD3- CD16+
CD56+ cells) in peripheral blood
Samples of venous blood were aliquoted into 12x75 mm plastic tubes (Falcon,
Becton Dickinson Company, USA), 100 µl per tube. Cells were stained with anti-CD3
ECD-labeled (Immunotech Beckman Coulter, France), anti-CD56 PE-labeled (BD PharMingen,
USA) and anti-CD16 FITC-labeled (BD PharMingen, USA) monoclonal antibodies (20
µg/test). For each set, an appropriate isotype control was prepared. After 30
min of incubation in the dark at room temperature, red blood cells were lysed
and samples fixed with the use of one-step Immunoprep reagent supplied by Immunotech,
USA with the use of Q-prep Immunology Workstation supplied by Coulter, USA.
The level of CD56 expression determines the phenotypic classification of NK
cells into CD56
dim NK cells exhibiting higher
natural cytotoxic activity and CD56
bright NK cells
revealing the capacity to produce cytokines and low natural cytotoxicity (19).
Determination of telomere length in NK cells by flow-FISH method
Telomeres were detected with the use of Telomere PNA Kit/FITC for Flow Cytometry
(DAKO, Denmark). Briefly, purified NK cells (1x10
6
cells) were suspended in 300 µl of hybridization mixture containing 70% formamide
with (test samples) or without (control samples) FITC-conjugated telomere PNA
(peptide nucleic acid) probe (CCCTAA)
3. Samples
were heat-denaturated at 82°C for 10 min and hybridised at room temperature
in the dark overnight. Then cells were washed twice with Wash Solution and stained
with propidium iodide solution (10 µg/ml propidium iodide and 100 µg/ml RN-ase
in PBS) for 2 hours. Flow cytometric analysis was performed on Coulter Epics
XL cytometer equipped with 488 nm argon laser. The obtained data were used for
the determination of a relative telomere length (RTL),
i.e. telomere
length of sample cells compared to telomere length of control cells (1301 cell
line characterized by very long telomeres) (20).
Estimation of telomerase activity in NK cells
Telomerase activity was measured in NK cell extracts with the use of TRAP
EZE®
ELISA Telomerase Detection Kit (Chemicon International, USA) following manufacturers
instructions. NK cells freshly isolated or activated with IL-2 (500 U/ml, 96
h) were suspended in lysis buffer containing CHAPS on ice for 30 min. After
centrifugation supernatants were transferred into fresh tubes, quick-frozen
in liquid nitrogen and stored in -70°C until use. TRAP (Telomeric Repeat Amplification
Protocol) reaction consisted of 2 steps: in the first one telomerase present
in extracts added a number of telomeric repeats (GGTTAG) to the 3 end of biotinylated
Telomerase Substrate oligonucleotide (b-TS) and in the second step the extended
products were amplified by PCR. As the TRAP extension/amplification reaction
was performed with biotinylated primers and DNP (dinitrophenyl)-labeled dCTP,
the TRAP products were tagged with biotin and DNP residues. The labeled products
were immobilized to streptavidin-coated microtiter plates
via biotin-streptavidin
interactions and then detected by anti-DNP antibodies conjugated to horseradish
peroxidase (HRP). The amount of TRAP products was determined by HRP activity
with the use of TMB substrate (3,3,5,5 tetramethylbenzidine) and subsequent
color development. Absorbance of the samples was measured at 450 nm (with a
reference wavelength of 690 nm) using BIORAD microplate reader.
Determination of intracellular IFN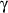 in NK cells
in NK cells
PBMC (1x10
6 cells/1 ml RPMI1640 containing 5%
FBS) were activated 5 h with PMA (phorbol 12-myristate 13-acetate) and ionomycin
both supplied by Sigma, USA and used at final concentration of 50 ng/ml. Simultaneously
2 µl/well of Golgi-Stop reagent (BD PharMingen, USA) was added to PBMC cultures
to stop extracellular export of IFN
.
Additionally, non-stimulated cultures were performed for the measurement of
spontaneous release of intracellular IFN
.
After 5 h PBMC were collected, washed with PBS (Ca
2+
and Mg
2+ free) containing 1% FBS and 0.09% sodium
azide and stained for subsequent 30 min with anti-CD3 ECD-labeled (Immunotech
Beckman Coulter, France) and anti-CD56 PE-labeled monoclonal antibodies (BD
PharMingen, USA). Then, cells were washed and fixed in 4% paraformaldehyde (Sigma,
USA) in PBS (Ca
2+ and Mg
2+
free), washed and permeabilised for 20 min using 0.1% saponin in PBS (Ca
2+
and Mg
2+ free) containing 1% FBS and 0.09% sodium
azide. After permeabilisation cells were washed, stained with anti-IFN
FITC-labeled monoclonal antibodies (1 µg/test) (BD PharMingen, USA) and incubated
for 30 min. Appropriate isotype controls were also prepared. Then samples were
washed, fixed in 2% paraformaldehyde and analyzed by flow cytometry with the
use of Coulter Epics XL cytometer equipped with 488 nm argon laser (21).
Statistics
Statistical analysis was performed with the use of STATISTICA 7.1 program. Parametric and non-parametric distributions were calculated by W Shapiro-Wilk test. As the analyzed data revealed no normal distribution they were then analyzed by non-parametric Kruskal-Wallis test to compare three age groups or U-Mann-Whitney test to compare two age groups. The experimental data are presented in the study as median and percentiles (10% / 90%).
RESULTS
Quantitative analysis of NK cells in the senior and young population
In our studies both young and two elderly groups did not show any differences
in the number of peripheral blood leukocytes. The old seniors (<85 ys), however,
showed a significant increase in the number of lymphocytes compared to the young
subjects (p = 0.0467). Percentage of lymphocytes as a fraction of all leukocytes
did not differ between the analyzed age groups (
Table 1).
| Table 1.
Percentages and absolute numbers of NK cells in peripheral blood of the
young, seniors under the age of ninety and seniors over the age of ninety.
The results are shown as median and percentiles (10% / 90%). Percentage
(%) is calculated for NK cells within lymphocyte gate and for lymphocytes
within leukocyte gate. Results statistically significant in the table
are marked with *. All differences among three age groups were calculated
by the Kruskal-Wallis test. All differences between two age groups were
calculated by the U-Mann-Whitney test (presented under the table only
for statistically significant differences, p < 0.05). |
 |
U-Mann Whitney
test:
Lymphocytes: young vs seniors < 85, p = 0.0467 (p < 0.05) |
The number of NK cells (CD3
-16
+56
+
cells) and the percentage of NK cells in the population of lymphocytes were
similar in all compared groups. Subsets of the CD56
+
cell population also did not display any significant differences between the
analyzed groups concerning both cell numbers and percentages in lymphocyte population
(
Table 1).
NK cytotoxic activity
Spontaneous NK cytotoxic activity in freshly isolated cells did not differ significantly
between the compared age groups although some age-dependent decrease was observed
in the old and the oldest subjects (
Fig. 1a). When PBMC from all age
groups were stimulated 5 h with PMA and ionomycin to estimate their ability
to produce intracellular IFN
it appeared that both the old (p=0.0329) and the oldest seniors (p=0.0051) showed
significant increase in the intracellular IFN
expression as compared to the young subjects (
Fig. 1b).
 |
| Fig. 1.
NK cytotoxic activity in the population of the young and seniors. A. NK
activity in the population of the young, old and the oldest. Data are
presented as median, 10th percentile / 90th percentile. B. Percentage
of NK cells with expression of intracellular IFN
(CD3-56+IFN+
cells) after 5 h of activation with PMA and ionomycin in the population
of the young, old and the oldest. Data are presented as median 10th percentile
/ 90th percentile (* p = 0.0329; # p = 0.0051). |
Telomere length and telomerase activity
Both telomere length and telomerase activity in NK cells decreased significantly
with age. Telomeres of both the old seniors (p=0.0007) and the oldest seniors
(p=0.0039) were significantly shorter than telomeres of the young persons. There
was, however, no difference between telomere length of the old and the oldest
subjects (
Fig. 2a).
Our studies revealed also that telomerase activity of the old was significantly
lower than activity of the young (p=0.0464) and telomerase activity of the oldest
was significantly lower than activity presented by the old (p=0.0444) (
Fig.
2b).
 |
| Fig. 2.
Telomere length and telomerase activity in NK cells of the young and senior
population. A. Telomere length in the population of the young, old and
the oldest presented in Relative Telomere Length units (RTL). Data are
shown as median 10th percentile / 90th percentile (* p = 0.0007; # p =
0.0039). B. Telomerase activity in NK cells isolated from the young, old
and the oldest (* p = 0.0464; # p = 0.0444) measured at 450 nm (A450)
presented in arbitrary units (AU). Data are shown as median 10th percentile
/ 90th percentile. |
Telomerase activity in resting and activated NK cells
After stimulation with IL-2 of NK cells isolated from PBMC of the young, old
and the oldest, all three age groups revealed an increase in telomerase activity.
No significant differences were observed between the analyzed groups but the
oldest seniors revealed tendency to present the highest increase in enzyme activity
relatively to control cells (
Fig. 3).
 |
| Fig. 3.
Telomerase activity in NK cells of the young, old and the oldest after
96 h of stimulation with IL-2 (500 U/ml) ) measured at 450 nm (A450)
presented in arbitrary units (AU). Control cells were cultivated simultaneously
for 96 h without stimulation. Data are presented as median 10th percentile
/ 90th percentile. |
DISCUSSION
Ageing of the immune system is characterized by the deterioration of both adaptive and innate immune response. Adaptive immune response is also affected by the proceeding process of thymic involution. Despite some age-related changes the innate immune system is considered to be less sensitive to ageing than adaptive response. Therefore it was of a great interest to compare one of the less investigated albeit important component of the innate immune response,
i.e. NK cells and their contribution to successful ageing.
The number of lymphocytes is a very general determinant of the ageing of the immune system, however, the literature data vary in this respect. Similarly to us also other authors (16, 22) found no differences between lymphocyte number of the young and elderly. However, there are studies which demonstrate that total number of lymphocytes may be increased in the elderly (23) or maintained at the middle-aged level up to the age of 95 and then decreased (24). It is thought that various factors including genetic ones may contribute to these discrepancies (17). The old population, however, presented in our studies a significant increase in the number of lymphocytes relatively to the young.
The studies related to the effect of ageing on the number/percentage of NK cells
reveal conflicting data. Similarly to other authors (25, 27) we found that the
oldest population of seniors did not differ significantly from the other age
groups in the number and percentage of NK cells and that there were no differences
both in cell numbers and percentages of CD56
+
subpopulations between the compared groups. Other authors found a significant
increase in the number of NK cells as well as an increase in the percentage
of more cytotoxic CD56
dim cells of the NK cell
population in aged people (25, 26). On the contrary, the number of CD56
bright
cells, the primary source of NK cell-derived IFN
appeared to decline with age (27). These differences may be partially attributed
to the health status of the elderly. Both the oldest and old subjects in our
study were generally healthy and had not shown any symptoms for longer time
before the investigation.
The cytotoxic activity of the NK cells is thought to be an important component of the ageing immune system. The NK activity of the oldest seniors in our study did not differ significantly from the other age groups. These data corresponded to the previous studies performed by Sansoni and coworkers who found that even very old seniors presented well preserved natural killer activity and this parameter could be interpreted as a factor of longevity (28). In earlier studies, however, NK cytotoxic activity was reported to be unaffected (29), decreased (30) or increased (31) in the elderly.
In normal physiology functioning of the immune system is continuously affected
by many activating and inhibiting factors that modulate its activity. Unexpectedly,
when we stimulated PBMC with nonspecific activators of lymphocytes, PMA and
ionomycin, we found that both groups of seniors displayed significantly higher
percentages of cells expressing intracellular IFN
as compared to young subjects. Moreover, we showed that NK cells of very old
seniors may effectively produce IFN
after antigenic stimulation. Interferon g strongly stimulates macrophages (32),
T
H1 lymphocytes (33) and cytotoxic T lymphocytes
(34). It also activates neutrophils and stimulates the cytotoxic activity of
NK cells (35). Our data are corroborated by study that showed significantly
higher rates of IFN
production in seniors attending moderate physical training program (36). The
ability of NK cells to produce IFN
can compensate age-related decrease in the adaptive immunity and help to preserve
the function of the immune system. However, some authors described diminished
secretion of IFN
after stimulation of NK cells of the elderly (8, 37). Again, the inherent differences
in the characteristics of the elderly subjects may be responsible for various
results between studies.
The length of telomeres is by some authors regarded as a major determinant of cellular ageing. The age-related shortening of telomeres of NK cells found in our investigation was also described by Mariani and coworkers (9). In their study only octogenarians revealed significantly shorter telomeres relatively to the septuagenarians and young, however, no significant differences were found in the telomere length of the young and septuagenarians (9). This observation with our finding of shorter telomeres (than in the young) in the old subjects (mean age 78 years) may suggest that there is an age threshold of telomere shortening in the NK cells. Further studies on a larger population could prove this suggestion.
The length of telomeres is to a great part dependent on the activity of telomerase. The oldest seniors in our study showed significantly lower NK cell telomerase activity than the old ones, whose NK cells had lower telomerase activity than the young subjects. Similar results related to septuagenarians and octogenarians obtained Mariani and coworkers (9). However, in their study, only octogenarians but not septuagenarians presented significantly lower telomerase activity than the young individuals.
Since IL-2 is a very potent immunomodulator acting on various lymphocyte populations we compared telomerase activity in NK cells isolated from the young, old and the oldest subjects. Interestingly, IL-2 increased telomerase activity to a similar range in the all investigated age groups. This is a novel observation as no similar investigations on NK cells isolated from healthy people in the context of ageing were performed. Kawauchi and coworkers described increase of telomerase activity in IL-2 stimulated cells of NK-92 cell line established from a patient with progressive NK cell lymphoma (38). It is known that telomerase activity is induced during activation of lymphoid cells (39) and its activity is probably important in determining the capacity of lymphoid cells to proliferation and clonal expansion (40).
In conclusion, we demonstrated that the NK component of the innate immunity is well preserved during ageing. The NK cells of very old and old subjects effectively respond to activation by increasing intracellular concentration of interferon g and an efficient increase of telomerase activity. The sensitivity of NK cells of nonagenarians to antigenic stimulation appears to contribute to healthy ageing and longevity.
Acknowledgements:
The authors thank Danuta Kazmierska for excellent technical support. The authors
are also greatly indebted to Professor Jacek M. Witkowski and Professor Zbigniew
Kmiec for critical comments and fruitful discussions.
Conflict of interest statement: None declared.
REFERENCES
- Cutolo M, Sulli A, Capellino S, et al. Sex hormones influence on the immune system: basic and clinical aspects in autoimmunity. Lupus 2004; 13: 635-638.
- Karasek M. Does melatonin play a role in aging processes? J Physiol Pharmacol 2007; 58 Suppl 6: 105-113.
- Szczepanik M. Melatonin and its influence on immune system. J Physiol Pharmacol 2007; 58 Suppl 6: 115-124.
- Sansoni P, Vescovini R, Fagnoni F, et al. The immune system in extreme longevity. Exp Gerontol 2008; 43: 61-65.
- Papamichail M, Perez SA, Gritzapis AD, Baxevanis CN. Natural killer lymphocytes: biology, development, and function. Cancer Immunol Immunother 2004; 53: 176-186.
- Mysliwska J, Bryl E, Chodnik T, Foerster J, Mysliwski A. Level of NK cytotoxic activity in the elderly aged more than 80 years. Arch Gerontol Geriatr 1992; 15: 21-28.
- Mysliwski A, Mysliwska J, Chodnik T, Bigda J, Bryl E, Foerster J. Elderly high NK responders are characterized by intensive proliferative response to PHA and Con A and optimal health status. Arch Gerontol Geriatr 1993; 16: 199-205.
- Mocchegiani E, Malavolta M. NK and NKT cell functions in immunosenescence. Aging Cell 2004; 3: 177-184.
- Mariani E, Meneghetti A, Formentini I, et al. Telomere length and telomerase activity: effect of ageing on human NK cells. Mech Ageing Dev 2003; 124: 403-408.
- Weng N. Interplay between telomere length and telomerase in human leukocyte differentiation and aging. J Leukoc Biol 2001; 70: 861-867.
- Proctor CJ, Kirkwood TBL. Modelling telomere shortening and the role of oxidative stress. Mech Ageing Dev 2002; 123: 351-363.
- Blackburn EH. Structure and function of telomeres. Nature 1991; 350: 569-572.
- Hornsby PJ. Telomerase and the aging process. Exp Gerontol 2007; 42: 575-581.
- Kim NW, Piatyszek MA, Prowse KR, et al. Specific association of human telomerase activity with immortal cells and cancer. Science 1994; 266: 2011-2015.
- Rowe JW, Kahn RL. Human aging: usual and successful. Science 1987; 237: 143-149.
- DelaRosa O, Pawelec G, Peralbo E, et al. Immunological biomarkers of ageing in man: changes in both innate and adaptive immunity are associated with health and longevity. Biogerontology 2006; 7: 471-481.
- Franceschi C, Monti D, Barbieri D, et al. Successful immunosenescence and the remodelling of immune responses with ageing. Nephrol Dial Transplant 1996; 11 (Suppl 9): 18-25.
- Olshansky SJ, Carnes BA, Cassel C. In search of Methuselah: estimating the upper limits to human longevity. Science 1990; 250: 634-640.
- Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends Immunol 2001; 22: 633-640.
- Larsson I, Lundgren E, Nilsson K, Strannegard O. A human neoplastic hematopoietic cell line producing a fibroblast type of interferon. Develop Biol Standard 1979; 42: 193-197.
- Hak L, Mysliwska J, Wieckiewicz J, et al. NK cell compartment in patients with coronary heart disease. Immun Ageing 2007; 4: 3.
- Pawelec G, Akbar A, Caruso C, Grubeck-Loebenstein B, Solana R, Wikby A. Human immunosenescence: is it infectious? Immunol Rev 2005; 205: 257-268.
- Herndler-Brandstetter D, Schwanninger A, Grubeck-Loebenstein B. CD4+ CD8+ T cells in young and elderly humans. Comment on Macchia I, Gauduin MC, Kaur A, Johnson RP. Expression of CD8a identifies a distinct subset of effector memory CD4+ T lymphocytes. Immunology 2006; 119: 232-42. Immunology 2007; 120: 292-294.
- Strindhall J, Nilsson BO, Lofgren S, et al. No immune risk profile among individuals who reach 100 years of age: findings from the Swedish NONA immune longitudinal study. Exp Gerontol 2007; 42: 753-761.
- Borrego F, Alonso MC, Galiani MD, et al. NK phenotypic markers and IL2 response in NK cells from elderly people. Exp Gerontol 1999; 34: 253-265.
- Solana R, Alonso MC, Pena J. Natural killer cells in healthy aging. Exp Gerontol 1999; 34: 435-443.
- Chidrawar SM, Khan N, Chan YLT, Nayak L, Moss PAH. Ageing is associated with a decline in peripheral blood CD56bright NK cells. Immun Ageing 2006; 3: 10.
- Sansoni P, Cossarizza A, Brianti V, et al. Lymphocyte subsets and natural killer cell activity in healthy old people and centenarians. Blood 1993; 82: 2767-2773.
- Bender BS, Chrest FJ, Adler WH. Phenotypic expression of natural killer cell associated membrane antigens and cytolytic function of peripheral blood cells from different aged humans. J Clin Lab Immunol 1986; 21: 31-36.
- Levy SM, Herberman RB, Lee J, et al. Persistently low natural killer cell activity, age, and environmental stress as predictors of infectious morbidity. Nat Immun Cell Growth Regul 1991; 10: 289-307.
- Krishnaraj R, Blandford G. Age-associated alterations in human natural killer cells. 1. Increased activity as per conventional and kinetic analysis. Clin Immunol Immunopathol 1987; 45: 268-285.
- Shenoy AR, Kim BH, Choi HP, Matsuzawa T, Tiwari S, MacMicking JD. Emerging themes in IFN-gamma-induced macrophage immunity by the p47 and p65 GTPase families. Immunobiology 2007; 212: 771-784.
- Kohno K, Kataoka J, Ohtsuki T, et al. IFN-gamma-inducing factor (IGIF) is a costimulatory factor on the activation of Th1 but not Th2 cells and everts its effect independently of IL-12. J Immunol 1997; 158: 1541-1550.
- Bose A, Baral R. IFNalpha2b stimulated release of IFNgamma differentially regulates T cell and NK cell mediated tumor cell cytotoxicity. Immunol Lett 2007; 108: 68-77.
- Gattoni A, Parlato A, Vangieri B, Bresciani M, Derna R. Interferon-gamma: biologic functions and HCV therapy (type I/II) (1 of 2 parts). Clin Ter 2006; 157: 377-386.
- Venjatraman JT, Fernandes G. Exercise, immunity and aging. Aging 1997; 9: 42-56.
- Solana R, Mariani E. NK and NK/T cells in human senescence. Vaccine 2000; 18: 1613-1620.
- Kawauchi K, Ihjima K, Yamada O. IL-2 increases human telomerase reverse transcriptase activity transcriptionally and posttranslationally through phosphatidylinositol 3-kinase/Akt, heat shock protein 90, and mammalian target of rapamycin in transformed NK cells. J Immunol 2005; 174: 5261-5269.
- Pan C, Xue BH, Ellis TM, Peace DJ, Diaz MO. Changes in telomerase activity and telomere length during human T lymphocyte senescence. Exp Cell Res 1997; 231: 346-353.
- Weng NP, Levine BL, June CH, Hodes RJ. Regulated expression of telomerase activity in human T lymphocyte development and activation. J Exp Med 1996; 183: 2471-2479.