Activation of group I mGluRs leads to a transient increase in intracellular calcium via the production of inositol-trisphosphates (3). Generally, it has been shown that activation of group I receptors enhances or facilitates the excitatory effects of glutamate by modulating ion channel activity (3). Antagonists of group I mGluRs have been proposed to exhibit potential therapeutic effects (3, 4) in such CNS disorders as epilepsy, ischemia, pain, depression and anxiety (3, 5-7).
Recent literature data and the results of our earlier pharmacological studies have provided evidence that the family of mGlu receptors play an important role in the anxiolytic-like properties of their ligands (6, 8-15). Involvement of group I mGluRs in anxiety has also been postulated. It has been shown that antagonists of group mGluR1 (which do not penetrate to brain) evoke anxiolytic-like effects after their intrahippocampal injection to rats (13), and that 2-methyl-6-(phenylethynyl)piridine (MPEP), a potent, non-competitive, brain-penetrating mGluR5 antagonist (16), administered systemically exhibits anxiolytic-like effects in several animal tests (12, 15, 17, 18). In the search for subtype-selective group I mGluR compounds, 1-aminoindan-1,5-dicarboxylic acid (AIDA) was synthesized and described a few years ago (19). AIDA was characterized as a selective, competitive mGluR1 antagonist, without any effects on group II or III mGluRs, or iGlu receptors (19-22). In the past few years, several reports on the electrophysiological, biochemical and pharmacological effects of AIDA after intracerebral (19, 21, 23-30) or intraperitoneal (31-33) administration have been published.
To the best of our knowledge, the effect of this drug has not yet been studied in animal models of anxiety; therefore we decided to examine its activity in the conflict drinking and plus-maze tests in rats, and in the four-plate test in mice - commonly used models for predicting a potential anxiolytic activity. Additionally, the effect of AIDA on the locomotor activity (open-field test) and motor coordination (rota-rod test) of rats was investigated. We used diazepam, a well-known anxiolytic, as a reference drug.
The doses of AIDA were chosen on the basis of studies by Nielsen et al. (31) and Christofferseen et al. (32), who observed significant effects of AIDA after itraperitoneal administration in behavioral experiments with rats.
General
Male Wistar rats (250±20g) and male Albino Swiss mice (25±2g) (Institute of Pharmacology PAS stock) were used for the study. The animals were kept in groups of six (rats) or thirty (mice) to a cage (53x36x18 cm) on a natural day-night cycle at a room temperature of 19-21°C, with free access to food and water. Each experimental group consisted of 6 - 10 animals/dose, and the animals were used only once for each test. The following groups were tested: controls injected with vehicle, and AIDA or diazepam treated animals (randomly chosen). Injection volume was 2 ml/kg for rats, and 10 ml/kg for mice. Experiments were carried out between 9:00 a.m. and 2:00 p.m. (from August until November). Behavioural responses were recorded by experimenter(s) unaware of the treatment the animals had received. All the experimental procedures were approved by the Animal Care and Use Committee at the Institute of Pharmacology of the Polish Academy of Sciences in Krakow.
Conflict drinking test (Vogel test) in rats
A modification of the method of Vogel et al. (34) was used. On the first day of the experiment, the rats were adapted to the test chamber for 10 min. It was a plexiglas box (27 x 27 x 50 cm), equipped with a grid floor made of stainless steel bars and a drinking bottle containing tap water. After the adaptation period, the animals were deprived of water for 24 h and were then placed in the test chamber for another 10-min adaptation period during which they had free access to the drinking bottle. Afterwards, they were allowed a 30-min free-drinking session in their home cage. After another 24-h water deprivation period, the rats were placed again in the test chamber and were allowed to drink for 30 s. Immediately afterwards, their drinking attempts were punished with an electric shock (0.5 mA). The impulses were released every 2 s (measured from the moment when the preceding shock was delivered) between the grid floor and the spout of the drinking bottle. Each shock lasted 1 s, and if a rat was drinking when an impulse was released, it received a shock. The number of shocks accepted throughout a 5-min experimental session was recorded.
Shock threshold and free-drinking tests in rats
To control for the possibility of occurrence of drug-induced changes in the perception of a stimulus or in the thirst drive which could contribute to behaviour in the conflict drinking test, stimulus threshold measurements and a free-drinking experiment were also carried out. In either case, the rats were treated before the experiment in the same way as in the conflict drinking test including two 24-h water deprivation periods separated by 30 min of water availability. In the shock threshold test, the rats were placed individually in the box, and electric shocks were delivered through the grid floor. The shock threshold was determined stepwise at 15 s shock-free intervals by increasing manually the current (0.1, 0.2, 0.3, 0.4 and 0.5 mA). A shock lasted 1 s and was delivered through the grid-floor until a rat showed an avoidance reaction (jump or jerk). In the free-drinking test, each animal was allowed to drink from the water spout. Licking was not punished. The total amount of water (ml) consumed during 5 min was recorded for each rat.
Elevated plus-maze test in rats
The construction and the testing procedure of an elevated plus-maze were based on a method described by Pellow and File (35). The wooden plus-maze apparatus, elevated to a height of 50 cm, consisted of two open arms (50x10 cm) and two closed arms (50x10 cm and 38 cm high walls), arranged so that the two arms of each type were opposite each other.
The plus-maze was placed in a darkened room and the center of the apparatus was illuminated with a 25 W electric bulb hanging 100 cm above. Each rat was placed in the centre of the plus-maze, facing one of the closed arms immediately after a 5-min adaptation in a wooden box (60x60x35 cm). During a 5-min test period, two experimenters who were sitting in the same room ca. 1 m from the end of one of the open arms recorded the number of entries into the closed or the open arms, as well as the time spent in either type of the arms. The entry with all four feet put onto one arm was defined as an arm entry. At the end of each trial the maze was wiped clean.
Four-plate test in mice
The box was made of an opaque plastic and had the shape of a rectangle (25x18x16 cm). The floor was covered with four rectangular metal plates (11x8 cm) separated by a gap of 4 mm. The plates were connected to a source of continuous current which enabled a 120V difference of potential between two adjacent plates for 0.5 s when the experimenter pressed a switch. Single mice were placed gently onto the plate, and were allowed to explore for 15 s. Afterwards, each time a mouse passed from one plate to another, the experimenter electrified the whole floor which evoked a visible flight reaction of the animal. If the animal continued running, it received no new shocks for the following 3 s. The number of punished crossings were counted for 60 s (36).
Open-field test in rats
The experiment was carried out on rats according to a slightly modified method by Janssen et al. (37). The center of the wooden open arena (100 cm in diameter), divided into six symmetrical sectors without walls, was illuminated with a 75 W electric bulb hanging directly 75 cm above it. Individual control or drug-injected animals were placed gently in the center of the arena and were allowed to explore freely. The time of walking, ambulation (the number of sector line crossings) and the number of rearing and peeping (looking under the edge of the arena) episodes were recorded by visual observation for 3 min.
Rota-rod test in rats
The rats were preselected 1 day before the test on the rotating rod (6 cm in diameter, 6 r.p.m.). The animals that held onto the rotating rod for 2 min were placed again on the same rotating rod on the following day, and were observed for 2 min. The number of animals falling off the rota-rod within 2 min was recorded.
Drugs
1-Aminoinoidan-1,5-dicarboxylic acid (AIDA, Tocris Cookson, Ltd., Bristol, UK) and diazepam (Polfa, Poznañ, Poland) were suspended in a 1% aqueous solution of Tween 80. Both substances or vehicle were administered intraperitoneally (i.p.) 60 min before the tests.
Analysis of the data
All the data were presented as the mean ± SEM. The statistical significance of the results was evaluated by a one-way ANOVA followed by Dunnett's Multiple Comparison Test. Rota-rod test data were calculated by Fisher's exact test.
Conflict drinking test in rats
AIDA, which was not effective in a dose of 0.5 mg/kg, in doses of 1 and 2 mg/kg significantly increased (by 243% and 217%, respectively) the number of shocks accepted during the experimental session in the Vogel test [F(3,24)=6.621, P<0.01; Figure 1]. Diazepam (used as a positive standard), administered in doses of 2.5, 5 and 10 mg/kg, increased (by 180, 373 and 466%, respectively) the number of the accepted shocks [F(3,26)=16.925, P<0.001; Figure 1].
 |
Fig. 1. Effects of AIDA and diazepam in the conflict drinking test in rats. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 6-8 rats per group) of the number of shocks accepted during a 5-min experimental session. * p<0.05, ** p<0.01 vs respective vehicle group. |
In control experiments, neither the threshold current nor water intake were changed by AIDA or diazepam administered in doses effective in the conflict drinking test, compared to the vehicle treatment (Table 1).
| Table 1. Effects of AIDA and diazepam on the shock threshold and the amount of water consumed by water-deprived rats. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 6 rats per group). |
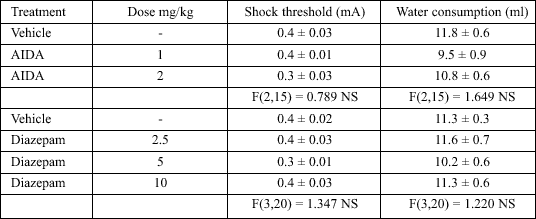 |
Plus-maze test in rats
The total number of entries (open + closed arm entries), observed in control rats during a 5-min test session, was about 6-7 in the present series of experiments and were taken as 100%. In control rats, 31.6 and 38.5% of the entries were made into the open arms (Figure 2); 10.7 and 10.9% of the total time (266 s) spent in the arms (either type) was spent in the open arms. AIDA administered in a dose of 0.25 mg/kg did not change the entries into and the time spent in the open arms. When given in doses of 0.5, 1 and 2 mg/kg, AIDA significantly increased the percentage of the time spent in the open arms up to 41, 55 and 49%, respectively [F(4,28)=6.231, P<0.01] and the percentage of entries into the open arms up to 50, 59 and 51%, respectively [F(4,28)=4.339, P<0.01] (Figure 2). AIDA given in doses 0.25, 0.5, 1 and 2 mg/kg did not change the entries into the closed arms (Table 2). AIDA in doses of 1 and 2 mg/kg significantly increased (by 76%) the total number of entries (data not shown). Diazepam, a positive standard, administered in a dose of 1.25 mg/kg, was ineffective in that test; however when given in doses of 2.5 and 5 mg/kg, it significantly increased the percentage of time spent in the open arms up to 47 and 70%, respectively [F(3,22)=14.52, P<0.001] as well as the percentage of entries into the open arms up to 74 and 76%, respectively [F(3,22)=5.871, P<0.01] (Figure 2). Diazepam given in doses of 1.25 and 2.5 mg/kg failed to affect the number of entries into the closed arms, given in a dose of 5 mg/kg significantly decreased the entries in the closed arms (Table 2). Diazepam in doses 5 mg/kg (but not lower) significantly reduced (by 52%) the total number of entries (data not shown).
 |
Fig. 2. Effects of AIDA and diazepam in the elevated plus-maze test in rats. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 6-7 rats per group). * p<0.05, ** p<0.01 vs respective vehicle group. |
| Table 2. Effects of AIDA and diazepam on the number of closed arm entries in the plus-maze test in rats. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 6 -7 rats per group). *p<0.05 vs respective vehicle group. |
 |
Four-plate test in mice
AIDA administered in doses of 0.5, 1, 2 and 8 mg/kg did not affect the number of mice's punished crossings in the four-plate test [F(4,45)=1.127, NS], while diazepam (1-4 mg/kg; used as a standard drug) increased in a dose-dependent manner the number of crossings [F(3,36)=12.807, P<0.001] (Figure 3).
 |
Fig. 3. Effects of AIDA and diazepam in the four-plate test in mice. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 10 mice per group). * p<0.05, ** p<0.01 vs respective vehicle group. |
Open field and rota-rod tests in rats
AIDA in the doses of 0.5-2 mg/kg did not change the exploratory locomotor activity of rats in the open field test, while its higher dose (4 mg/kg) significantly increased the time of walking and ambulation in that species (Table 3). Diazepam in a dose of 2.5 mg/kg did not change the exploratory locomotor activity of rats in the open field test, whereas given in doses of 5 and 10 mg/kg significantly decreased all the parameters measured (Table 3). AIDA in doses up to 4 mg/kg did not disturb the motor coordination of rats on the rotating rod (data not shown; P=1.0000, NS).
| Table 3. Effects of AIDA and diazepam on the exploratory activity of rats in the open field test. AIDA and diazepam were administered 60 min before the test. The given values represent the mean ± SEM (n = 6 rats per group). *p<0.05, **p<0.01 vs respective vehicle group. |
 |
In order to further investigate the role of group I mGluRs receptors in anxiety, we evaluated the action of AIDA, a selective mGluR1 antagonist (19, 21), in three animal models of anxiety. The obtained results showed that - like diazepam, used as a reference drug - AIDA exerted potential anxiolytic activity in rats. In fact, the drug tested increased the number of shocks accepted in the conflict drinking Vogel test. The action of AIDA seems to be specific, since this drug in doses effective in the Vogel test affects neither the response to threshold current nor water intake. The Vogel test is a widely employed model, and is considered to by one of the most specific methods for the detection of potential anxiolytic activity (38). The results obtained in the Vogel test are supported by the anxiolytic-like effect of AIDA detected by the elevated plus-maze test, a procedure based on a rodent's natural aversion to height and open space. AIDA increased the percentage of time spent in and the number of entries into the open arms of the plus-maze, whereas it had no significant effect on the number of entries into the closed arms. The total number of entries (open + closed arm entries/test session) was taken as an index of drug effect on locomotor activity (39). AIDA caused a small but significant increase in the total number of entries into the arms of the maze; on the other hand, when administered in doses effective in the elevated plus-maze test, it did not change the exploratory activity of rats measured in the open field test. Therefore the AIDA-induced increase in the percentage of open arm entries and time spent in the open arms seems to reflect a specific antianxiety effect, and cannot be explained by competing behaviour such as, e.g., general locomotor activity.
The bell-shaped dose response curve, observed in our experiment in animal anxiety models, is characteristic of a number of drugs and substances with anxiolytic properties, such as, e.g., pentobarbital (34), buspirone (40), L-701,324 an antagonist at glycineB receptors (41) and LY 354740, a group II mGluR agonist (11). The anxiolytic-like activity of AIDA is in line with several observations that other mGluR1 antagonists, i.e. (s)-4-CPG ((S)-4-carboxyphenylglycine) and CPCCOEt (7-(hydroxyimino) cyclopropan [b]chromen-1alpha-carboxylic ethyl ester), exert a similar action after their intrahippocampal injection (13). Moreover, as outlined briefly in the introduction we previously reported that MPEP, a selective antagonist of mGluR5, produced anxiolytic-like activity in the Vogel test and the elevated plus-maze test in rats, and in the four-plate test in mice (12). An anxiolytic-like effect of MPEP was also demonstrated in a social interaction test and the stress-induced hypothermia (15, 18). In the four-plate test in mice i.e., the only test involving mice, AIDA (in contrast to diazepam) failed to exert anxiolytic-like activity, even when it was used in doses several times higher than those inducing positive effects in the Vogel or the plus-maze tests. It is noteworthy that in the four-plate test MPEP in a high dose (30 mg/kg) produced only a weak anxiolytic-like effect, whereas in other models it showed such activity in tenfold lower doses.
Interestingly enough, while benzodiazepine receptor agonists have been shown to produce anxiolytic-like activity in the majority of anxiety models, non-benzodiazepine compounds showing some efficacy in the treatment of anxiety disorders, or proposed as anti-anxiety agents have yielded inconsistent results in these procedures (42). The reason for such discrepancies is not fully understood, but obviously many factors can be mentioned here, eg. the procedures used, the level of stress, or the species of animals and the resulting differences in brain penetration and receptor occupancy. For example, recently Anderson et al. (43) showed differences in in vivo receptor occupancy in rats and mice after treatment with mGluR5 antagonists. At the same time, those authors observed that compound plasma levels were substantially lower in mice than in rats, which suggested that species differences in receptor occupancy were probably due to variations in the absorbtion or metabolism of compounds. Therefore it cannot be to excluded that the lack of the anxiolytic-like activity of AIDA in a four-plate test in mice is a consequence of its weaker receptor occupancy, induced by more rapid metabolism and/or reduced absorbtion in that animal species.
The results of the experiments described above indicate that antagonists of mGluR1 - like those of mGluR5 - may have a benefitial effect in anxiety. The mechanism of such an effect may by related to the reduction of excitatory glutamatergic neurotransmission. Antagonists of mGluR1 and mGluR5 (either belonging to group I mGluRs) may simply counteract the excitation mediated by stimulation of these receptor types (3, 44). It has been shown that both mGlu1 and mGlu5 receptors are expressed in rat hippocampus (45-49), one of the brain structures involved in anxiety (50) and anti-anxiety effects of different compounds modulating the function of the glutamatergic system (6) including antagonists of mGluR1 (13). The decreased glutamatergic transmission, which leads to overall inhibitory effects in the CNS, may have consequences similar to the effect of the increased GABA-ergic transmission, considered to underlie the anxiolytic effects of benzodiazepines (51). The AIDA-induced increase in the hippocampal output of GABA (25) should be taken into account as a possible mechanism of its anxiolytic-like effects.
Although there have been no available data on the brain concentration of AIDA after its peripheral administration, the central effects described by these and other authors (31-33) do appear after such administration of the drug. Therefore it remains to be elucidated whether the observed effects of AIDA in vivo (after peripheral administration) stem - via the blockade of mGluR1 receptors - from the drug's own action, or/and are rather prompted by some active AIDA metabolites, not described so far; hence involvement of other receptors cannot be totally excluded.
In conclusion, AIDA, a selective antagonist of mGluR1, was found to produce anxiolytic-like effects in the Vogel and the plus-maze tests in rats. It was also observed that AIDA - in contrast with diazepam - neither induced sedation nor disturbed motor coordination in animals. Since the treatment of anxiety requires long-term drug administration, it seems necessary to determine whether tolerance to and/or dependence on the anxiolytic-like effect of AIDA develops after prolonged treatment.
- Monaghan DT, Bridges RJ, Cotman CW. The excitatory amino acid receptors: their classes, pharmacology, and distinct properties in the function of the central nervous system. Ann Rev Pharmacol Toxicol 1989; 29: 365-402.
- Pin JP, Duvoisin R. The metabotropic glutamate receptors: structure and functions. Neuropharmacology 1995; 34: 1-26.
- Conn PJ, Pin JP. Pharmacology and functions of metabotropic glutamate receptors. Ann Rev Pharmacol Toxicol 1997; 37: 205-237.
- Bruno V, Copani A, Battaglia G, Raffaele R, Shinozaki H, Nicoletti F. Protective effect of the metabotropic glutamate receptor agonist, DCG-IV, against excitotoxic neuronal death. Eur J Pharmacol 1994; 256:109-112.
- Nicoletti F, Bruno V, Copani A, Casabona G, Knopfel T. Metabotropic glutamate receptors: A new target for the therapy of neurodegenerative disorders? Trends Neurosci 1996; 19: 267-271.
- Chojnacka-Wojcik E, Klodzinska A, Pilc A. Glutamate receptor ligands as anxiolytics. Curr Opin Invest Drugs 2001; 2: 1112-1119.
- Pilc A, Klodzinska A, Nowak G. A role for glutamate in the treatment of anxiety and depression: focus on group I metabotropic glutamate (mGlu) receptor. Drugs of the Future 2002; 27: 1-11.
- Chojnacka-Wojcik E, Tatarczynska E, Pilc A. The anxiolytic-like effect of metabotropic glutamate receptor antagonists after intrahippocampal injection in rats. Eur J Pharmacol 1997; 319: 153-156.
- Monn JA, Valli MJ, Massey SM et al. Design, synthesis, and pharmacological characterization of (+)-2-aminobicyclo [3.1.0.]hexane-2,6-dicarboxylic acid (LY354740): a potent, selective, and orally active group 2 metabotropic glutamate receptor agonist possessing anticonvulsant and anxiolytic properties. J Med Chem 1997; 40: 528-537.
- Helton DR, Tizzano JP, Monn JA, Schoepp DD, Kallman MJ. Anxiolytic and side-effect profile of LY 354740: a potent, highly selective, orally active agonist for group II metabotropic glutamate receptors. J Pharmacol Exp Ther 1998; 284: 651-660.
- Klodzinska A, Chojnacka-Wojcik E, Palucha A, Branski P, Popik P, Pilc A. Potential anti-anxiety, anti-addictive effects of LY 354740, a selective group II glutamate metabotropic receptors agonist in animal models. Neuropharmacology 1999; 38: 1831-1839.
- Tatarczynska E, Klodzinska A, Chojnacka-Wojcik E et al. Potential anxiolytic- and antidepressant-like effects of MPEP, a potent, selective and systemically active mGlu5 receptor antagonist. Br J Pharmacol 2001; 132: 1423-1430.
- Tatarczynska E, Klodzinska A, Kroczka B, Chojnacka-Wojcik E, Pilc A. The antianxiety-like effects of antagonists of group I and agonists of group II and III metabotropic glutamate receptors after intrahippocampal administration. Psychopharmacology 2001; 158: 94-99.
- Pilc A, Klodzinska A, Branski P et al. Multiple MPEP administrations evoke anxiolytic- and antidepressant-like effects in rats. Neuropharmacology 2002; 43: 181-187.
- Spooren WPJM, Schoeffter P, Gasparini F, Kuhn R, Gentsch K. Pharmacological and endocrinological characterisation of stress-induced hyperthermia in singly housed mice using classical and candidate anxiolytics (LY 314582, MPEP and NKP608). Eur J Pharmacol 2002; 435: 161-170.
- Gasparini F, Lingenhohl K, Stoehr N et al. 2-methyl-6-(phenylethynyl)-pyridine (MPEP), a potent, selective and systemically active mGlu5 receptor antagonist. Neuropharmacology 1999; 38: 1493-1503.
- Klodzinska A, Tatarczynska E, Chojnacka-Wojcik E, Pilc A. Anxiolytic-like effects of group I metabotropic glutamate antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP) in rats. Pol J Pharmacol 2000; 52: 463-466.
- Spooren WPJM, Vassout A, Neijt HC et al. Anxiolytic-like effects of the prototypical metabotropic glutamate receptor 5 antagonist 2-methyl-6-(phenylethynyl)pyridine in rodents. J Pharmacol Exp Ther 2000; 295: 1267-1275.
- Pellicciari R, Luneia R, Costantino G et al. 1-Aminoindan-1,5-dicarboxylic acid: a novel antagonist at phospholipase C-linked metabotropic glutamate receptors. J Med Chem 1995; 38: 3717-3719.
- Costantino G, Pellicciari R. Homology modeling of metabotropic glutamate receptors (mGluRs): structural motifs affecting binding modes and pharmacological profile of mGluR1 agonist and competitive antagonists. J Med Chem 1996; 39: 3998-4006.
- Moroni F, Lombardi G, Thomsen C et al. Pharmacological characterization of 1-aminoindan-1,5-dicarboxylic acid, a potent mGluR1 antagonist. J Pharmacol Exp Ther 1997; 281: 721-729.
- Hermans E, Challiss RAJ, Nahorski SR. Effects of varying the expression level of recombinant human mGlu1a receptors on the pharmacological properties of agonists and antagonist. Br J Pharmacol 1999; 126: 873-882.
- Thomsen C, Dalby NO. Roles of metabotropic glutamate receptor subtypes in modulation of pentylenetetrazole-induced seizure activity in mice. Neuropharmacology 1998; 37: 1465-1473.
- Chapman AG, Yip PK, Yap JS et al. Anticonvulsant actions of LY 367385 ((+)-2-methyl-4-carboxyphenylglycine) and AIDA ((RS)-1-aminoindan-1,5-dicarboxylic acid). Eur J Pharmacol 1999; 368: 17-24.
- Pellegrini-Giampietro DE, Peruginelli F, Meli E et al. Protection with metabotropic glutamate 1 receptor antagonists in models of ischemic neuronal death: time-course and mechanisms. Neuropharmacology 1999; 38: 1607-1619.
- Mills CD, Xu GY, Johnson KM, McAdoo DJ, Hulsebosch CE. AIDA reduces glutamate release and attenuates mechanical allodynia after spinal cord injury. Neuroreport 2000; 11: 3067-3070.
- Wolfarth S, Konieczny J, Lorenc-Koci E, Ossowska K, Pilc A. The role of metabotropic glutamate receptor (mGluR) ligands in parkinsonian muscle rigidity. Amino Acids 2000; 19: 95-101.
- Car H, Nadlewska A, Oksztel R, Winiewski K. AIDA influences behavior in rats pretreated with baclofen. Pol J Pharmacol 2001; 53: 245-252.
- Cozzi A, Meli E, Carla V, Pellicciari R, Moroni F, Pellegrini-Giampietro DE. Metabotropic glutamate 1 (mGlu1) receptor antagonists enhance GABAergic neuro-transmission: a mechanism for the attenuation of post-ischemic injury and epileptiform activity? Neuropharmacology 2002; 43: 119-130.
- Dolan S, Nolan AM. Behavioral evidence supporting a differential role for spinal group I and II metabotropic glutamate receptors in inflammatory hyperalgesia in sheep. Neuropharmacology 2002; 43: 319-326.
- Nielsen KS, Macphail EM, Riedel G. Class I mGlu receptor antagonist 1-aminoindan-1,5-dicarboxylic acid blocks contextual but not cue conditioning in rats. Eur J Pharmacol 1997; 326: 105-108.
- Christoffersen GRJ, Christensen LH, Harrington NR, Macphail EM, Riedel G. Task-specific enhancement of short-term, but not long-term, memory by class I metabotropic glutamate receptor antagonist 1-aminoindan-1,5-dicarboxylic acid in rats. Behav Brain Res 1999; 101: 215-226.
- Rao AM, Hatcher JF, Dempsey RJ. Neuroprotection by group I metabotropic glutamate receptor antagonists in forebrain ischemia of gerbil. Neurosci Lett 2000; 293: 1-4.
- Vogel JR, Beer B, Clody DE. A simple and reliable conflict procedure for testing anti-anxiety agents. Psychopharmacologia 1971; 21: 1-7.
- Pellow S, File SE. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacol Biochem Behav 1986; 24: 525-529.
- Aron C, Simon P, Larousse C, Boissier JR. Evaluation of a rapid technique for detecting minor tranquilizers. Neuropharmacology 1971; 10: 459-469.
- Janssen PA, Jageneau AH, Schellekens KH. Chemistry and pharmacology of compounds related to 4-(4-hydroxy-4-phenyl-piperidino)-butyrophenone. IV. Influence of haloperidol (R 1625) and of chlorpromazine on the behaviour of rats in an unfamiliar 'open field' situation. Psychopharmacologia 1960; 1: 389-392.
- Pollard GT, Howard JL. Effects of drugs on punished behavior: pre-clinical test for anxiolytics. Pharmacol Ther 1990; 45: 403-424.
- Dawson GR, Tricklebank MD. Use of the elevated plus-maze in the search for novel anxiolytic agents. Trends Pharmacol Sci 1995; 16: 33-36.
- Przegalinski E, Tatarczynska E, Chojnacka-Wojcik E. Anticonflict effect of ipsapirone, buspirone and gepirone is not mediated by their common metaboilte 1-(2-pyrimidinyl)piperazine. J Psychopharmacol 1989; 3: 180-185.
- Przegalinski E, Tatarczynska E, Chojnacka-Wojcik E. Anxiolytic- and antidepressant- like effects of an antagonist at glycineB receptors. Pol J Pharmacol 1998; 50: 349-354.
- Belzung C. Rodent models of anxiety-like behaviors: Are they predictive for compounds acting via non-benzodiazepine mechanisms? Curr Opin Invest Drugs 2001; 2: 1108-1111.
- Anderson JJ, Bradbury MJ, Giracello DR, et al. In vivo receptor occupancy of mGlu5 receptor antagonists using the novel radioligand [3H]3-methoxy-5-(pyridin-2-ylethynyl)pyridine). Eur J Pharmacol 2003; 473: 35-40.
- Bordi F, Ugolini A. Group I metabotropic glutamate receptors: implications for brain diseases. Prog Neurobiol 1999; 59: 55-79.
- Masu M, Tanabe Y, Tsuchida K, Shigemoto R, Nakanishi S. Sequence and expression of a metabotropic glutamate receptor. Nature 1991; 349: 760-765.
- Abe T, Sugihara H, Nawa H, Shigemoto R, Mizuno N, Nakanishi S. Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca2+ signal transoluction. J Biol Chem 1992; 267: 13361-13368.
- Martin LJ, Blackstone CD, Huganir RL, Price DL. Cellular localization of a metabotropic glutamate receptor in rat brain. Neuron 1992; 9: 259-270.
- Shigemoto R, Nomura S, Ohishi H, Sugihara H, Nakanishi S, Mizuno N. Immunohistochemical localization of a metabotropic glutamate receptor mGluR5, in the rat brain. Neurosci Lett 1993; 163: 53-57.
- Testa CM, Friberg IK, Weiss SW, Standaert DG. Immunohistochemical localization of metabotropic glutamate receptors mGluR1a and mGlu2/3 in the rat basal ganglia. J Comp Neurol 1998; 390: 5-19.
- Gray JA. Precis of the neuropsychology of anxiety: an enquiry into the functions of the septo-hippocampal system. Behav Brain Sci 1982; 5: 469-534.
- Haefely W, Kulcsar A, Mohler H, Pieri L, Polc P, Schaffner R. Possible involvement of GABA in the central actions of benzodiazepines. Adv Biochem Psychopharmacol 1975; 14: 131-151.