It is commonly accepted that sigma receptor ligands can modulate neurotransmission mediated by central neurotransmitter systems, including dopaminergic and glutamatergic/NMDA ones, which are seemingly important for the mechanism of action of ADs (10-12). The antidepressive properties of NMDA receptor antagonists were postulated more than ten year ago and their antidepressant-like action was shown in a number of animal models (e.g. 11, 13-15). Our earlier studies showed that ADs (imipramine, citalopram, mianserin) potentiated the antidepressant-like effect of MK-801, uncompetitive NMDA receptor antagonist, indicating the existence of synergism between ADs and MK-801 (13). Moreover, ADs administered in combination with amantadine, memantine or neramexan, uncompetitive NMDA receptor antagonists, display a positive interaction in the forced swimming test in rats. Therefore it is possible that co-administration of ADs and NMDA receptor antagonists may induce a more pronounced antidepressive activity in clinic than treatment with ADs alone (16). The latter observation may be of particular importance in the case of drug-resistant patients and may suggest a method of obtaining significant antidepressive actions with limited side-effects.
Our preliminary results indicated that selective sigma1 and sigma2 receptor agonists given jointly with AMA decreased the immobility time of rats (8). The present study was aimed at determining whether combined treatment with SA4503 or siramesine on the one hand and MEM (another uncompetitive NMDA receptor antagonist) on the other, led to an increase in antidepressant-like activity compared to the effects of those compounds given separately. Additionally, we used progesterone, a sigma1 receptor antagonistic neurosteroid, and BD 1047, a novel sigma antagonist with preferential affinity for sigma1 sites (17, 18), to determine the role of the sigma1 receptor in the effect induced by joint treatment with sigma ligands and the uncompetitive NMDA receptor antagonists - AMA and MEM.
Animals and drug treatment
The experiments were carried out on rats (male Wistar, 250-300 g) housed in groups (6 per cage) in a controlled environment at a temperature of 22±2°C on a 12- hour light/dark cycle (the light on at 7 a.m.). The animals had free access to food and water. AMA, MEM, SA4503, BD 1047 and sulpiride were dissolved in distilled water (sulpiride with a slight addition of citric acid), progesterone in a 10% DMSO + a 1% aqueous solution of carboxymethylcellulose; siramesine and prazosin were suspended in a 1% aqueous solution of Tween 80. The compounds were administered i.p. (AMA, MEM, BD 1047, prazosin, siramesine, sulpiride), s.c. (progesterone) or p.o. (SA 4503) at a volume of 2 ml/kg. Studies were conducted between 8 a.m. and 3 p.m. Experimental protocols were approved by the Local Bioethics Commission and to comply with the guidelines of the agency in charge at the Institute of Pharmacology, Polish Academy of Sciences.
Drugs
Amantadine hydrochloride (AMA, Sigma-Aldrich Chemie GmbH, Germany), BD 1047 (N-[2-(3,4-dichlorophenyl)ethyl]-N-methyl-2-(dimethylamino) ethylamine, Tocris, UK), memantine hydrochloride (MEM, Sigma-Aldrich Chemie GmbH, Germany), prazosin (hydrochloride, Research Biochemicals Inc., USA), progesterone (4-pregnene 3, 20-dione, Serva Feinbiochemica, Germany), SA4503 (1-(3,4-dimethoxyphenethyl)-4-(3-phenylpropyl)piperazine dihydrochloride, Santen Pharmaceutical Co. Ltd., Japan), siramesine hydrochloride (Lundbeck, Denmark), (-) sulpiride (Research Biochemicals Inc., USA).
Forced swimming (Porsolt's) test
The animals were subjected to two trials during which they were forced to swim in a cylinder (40 cm high, 18 cm in diameter) filled with water (23-25°C) up to a height of 15 cm. There was a 24-hour interval between the first and the second trial; the first trial lasted 15 min, the second 5 min. The total duration (s) of immobility was measured throughout the second trial (19).
SA4503 (3 mg/kg) and siramesine (1 mg/kg) were given separately or in combination with AMA (10 mg/kg) or MEM (2.5 mg/kg) three times: at 24, 5, and 1 h before the test. In some experiments, progesterone (20 mg/kg) or BD1047 (3 mg/kg) was given at 15 min, sulpiride or prazosin - at 30 min before joint treatment with sigma ligands and NMDA antagonists. Each group consisted of 8 rats.
Locomotor activity test
The locomotor activity of rats was studied with photoresistor actometers (two light beams; LxWxH = 40x40x25 cm) for 30 min. Drug treatments were carried out according to the same experimental schedule as described above (three injections). Each group consisted of 6-8 rats.
Statistical analysis
The data were evaluated by an analysis of variance (ANOVA), followed - when appropriate - by individual comparisons with the control using Dunnett's test.
Forced swim (Porsolt's) test
SA4503 at the doses used (1, 3 and 10 mg/kg) did not modify the immobility time of rats in a statistically significant manner. Co-administration of SA4503 and MEM (in a dose of 2.5 mg/kg, inactive per se) induced a decrease in the immobility time (Fig 1A). Siramesine, 1 and 3 mg/kg, as well as MEM, did not influence the immobility time of rats. Joint treatment with siramesine and MEM produced an antidepressant-like effect (Fig. 1B).
 |
Fig. 1. The effect of combined treatment with memantine (MEM) and SA4503 (A) or siramesine (SIR) (B) in the forced swimming test in rats. MEM (2.5 mg/kg) and SA4503 (1, 3, 10 mg/kg) or SIR (1, 3 mg/kg) were given three times at 24, 5 and 1 h before the test. The obtained results represent the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
SA4503 (3 mg/kg) administered in combination with AMA (10 mg/kg) decreased the immobility time of rats compared to the control (vehicle) group, as well as to SA4503 (3 mg/kg) or AMA (10 mg/kg) group. Progesterone (20 mg/kg) completely abolished that effect (Fig. 2).
 |
Fig. 2. The influence of progesterone on the anti-immobility effect of SA4503 and amantadine (AMA). SA4503 (3 mg/kg) or AMA (10 mg/kg), or both compounds jointly were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with progesterone (20 mg/kg; 15 min before each injection of SA4503, AMA, or their co-administration). The obtained results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
Co-admnistration of SA4503 (3 mg/kg) and MEM (2.5 mg/kg) induced an antidepressant-like effect, which was partly antagonized by progesterone (20 mg/kg) (Fig. 3).
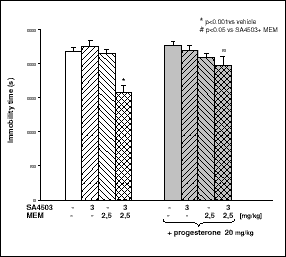 |
Fig. 3. The influence of progesterone on the anti-immobility effect of SA4503 and MEM. SA4503 (3 mg/kg) or MEM (2.5 mg/kg), or both compounds jointly were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with progesterone (20 mg/kg; 15 min before each injection of SA4503, MEM, or their co-administration). The obtained results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
Siramesine (1 mg/kg) given jointly with AMA (10 mg/kg) shortened the time of immobility. Progesterone (20 mg/kg) did not modify that effect (Fig. 4). Similar lack of effect was observed when the rats were pretreated with BD 1047 (3 mg/kg) (Fig. 5A). The antidepressant-like effect evoked by joint treatment with siramesine and AMA was partly counteracted by sulpiride (10 mg/kg) (Fig. 5B) and prazosin (1 mg/kg) (Fig. 5C). The anti-immobility effect induced by co-administration of siramesine and MEM was not changed by progesterone (Fig. 6) but attenuated by BD 1047 (Fig. 7A), sulpiride (Fig. 7B) and prazosin (Fig. 7C).
 |
Fig. 4. The influence of progesterone on the anti-immobility effect of SIR and AMA. SIR (1 mg/kg) or AMA (10 mg/kg), or both compounds jointly were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with progesterone (20 mg/kg; 15 min before each injection of SIR, AMA, or their co-administration). The results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
 |
Fig. 5. The influence of BD 1047 (A), sulpiride (B), or prazosin (C) on the anti-immobility effect of SIR and AMA. SIR (1 mg/kg) and AMA (10 mg/kg) were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with BD 1047 (A) (3 mg/kg; 15 min before each injection of SA4503, AMA, or their co-administration), sulpiride (B) (10 mg/kg, 30 min before each injection of SA4503, AMA, or their co-administration), or prazosin (C) (1 mg/kg, 30 min before each injection of SIR , AMA, or their co-administration). The obtained results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
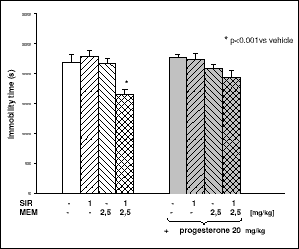 |
Fig. 6. The influence of progesterone on the anti-immobility effect of SIR and MEM. SIR (1 mg/kg) or MEM (2.5 mg/kg), or both compounds jointly were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with progesterone (20 mg/kg; 15 min before each injection of SA4503, MEM, or their co-administration). The obtained results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
 |
Fig. 7. The influence of BD 1047 (A), sulpiride (B), or prazosin (C) on the anti-immobility effect of SIR and MEM. SIR (1 mg/kg) or MEM (2.5 mg/kg), or both compounds jointly were given three times at 24, 5 and 1 h before the test. The right side of the columns (gray background) represents groups pretreated with BD 1047 (A) (3 mg/kg; 15 min before each injection of SIR, MEM, or their co-administration), sulpiride (B) (10 mg/kg; 30 min before each injection of SIR, MEM, or their co-administration), or prazosin (C) (1 mg/kg; 30 min before each injection of SIR, MEM, or their co-administration). The obtained results show the mean ± SEM; n=8. The data were statistically evaluated by ANOVA, followed by individual comparisons using Dunnett's test. |
The locomotor activity test in rats
The sigma agonists, SA4503 (3 mg/kg) and siramesine (1 mg/kg), or the uncompetitive NMDA receptor antagonists AMA (10 mg/kg) and MEM (2.5 mg/kg), or their combinations did not modify the locomotor activity of rats (data not shown).
Although the mechanism of the action of ADs has been studied for many years, it still remains unclear in details and successive new hypotheses are proposed. In accordance with the present opinion, the third generation of ADs should activate monoaminergic transmission and increase the corticolimbic availability of monoamines through indirect mechanisms or by acting "downstream", by cellular substrates controlling gene expression, synaptic plasticity and neurogenesis (20). Such conditions are partly satisfied by selective sigma ligands, which are able to indirectly modulate monoaminergic transmission (21).
As has been proposed by Hayashi et al. (22), sigma receptors control the intracellular levels of Ca2+. Moreover, they can also modulate ion channels, e.g. voltage-dependent Ca2+ channels, as well as NMDA receptor and K+ channels (the former by direct coupling as a modulatory subunit) (23). Such mechanisms, being related to a downstream target, may be involved in the potential antidepressant effect of selective sigma ligands.
On the basis of biochemical in vitro experiments, SA4503 was characterized as a sigma1 receptor agonist (24). Preliminary studies indicated that it decreased immobility time in the tail suspension test and Porsolt's test in mice (7, 9). Alleviation of behavioral despair in mice was also observed after another sigma1 agonist, (+)pentazocine, and after the sigma1/sigma2 agonist 1,3di-o-tolylguanidine (DTG). Moreover, those antidepressant-like effects were antagonized by the sigma1 antagonist NE-100 (7), or by panamesine, a potential antipsychotic drug with non-selective sigma receptor antagonistic activity (25). The antidepressant-like effect of other sigma1 agonists, igmesine (JO 1783) and OPC 14523, was recently demonstrated in preclinical studies (26, 27).
However, sigma2 receptors are also likely to modulate emotional responses, since siramesine (a selective sigma2 agonist) showed an anxiolytic-like effect in black-and-white two-compartment box tests in mice and rats, in a social interaction test in rats and in the Vogel conflict test. Moreover, siramesine had moderate antidepressant activity in a chronic mild stress (CMS) model (28, 29). It is noteworthy that no selective antagonists of the sigma2 receptor have been known so far.
Interactions between neuroactive steroids and the sigma1 receptor have been described in in vitro and in vivo studies (for review see: 30, 31). It was shown that dehydroepiandrosterone (DHEA), pregnenolone or their sulfate esters behaved as agonists of the sigma1 receptor, whereas progesterone - as its antagonist. In the forced swimming test, DHEA sulfate and pregnenolone sulfate caused reduction of immobility, which was reversed by the selective sigma1 receptor antagonist NE-100. On the other hand, progesterone blocked both the sigma1 receptor agonist-mediated effect and neurosteroid effects. A similar cross-reaction between sigma1 receptor ligands and neuroactive steroids was observed at the behavioral level, e.g. in several amnesia models (30, 31).
The anti-immobility effect of SA4503 in the Porsolt's test in rats is not pronounced. It is important to know that a biphasic, bell-shaped dose-response curve has been observed for sigma ligands in various behavioral, biochemical and electrophysiological paradigms (32). In the present study, SA4503 did not decrease immobility time in a statistically significant manner (at the doses used). However, when given jointly with AMA or MEM, it produced antidepressant-like activity. Siramesine per se failed to show any activity in Porsolt's test in rats. Nevertheless, its co-administration with AMA or MEM caused a decrease in the immobility time of rats, but only at one dose tested (the same as that active in the CMS test) (8). The antidepressant-like effect of combined treatment with SA4503 and AMA was completely antagonized by progesterone. Likewise, progesterone attenuated the anti-immobility effect of joint treatment with SA4503 and MEM.
In contrast, neither progesterone nor BD1047 counteracted the effect of combined treatment with siramesine and AMA, whereas the antidepressant-like effect of the latter compounds was attenuated by sulpiride and prazosin (a dopamine D2 - and an alpha1-adrenergic receptor antagonists, respectively). The lack of effect of progesterone and BD1047 seems to be justifiable, considering the sigma1 antagonistic activity of both these compounds. However, besides its high affinity for sigma1 sites (Ki = 0.93 nM), BD1047 also binds to the sigma2 receptor (Ki = 47nM ) (17). The results obtained in the present study suggest that the anti-immobility effect of combined treatment with siramesine and AMA or MEM is related to noradrenergic and dopaminergic mechanisms, but not to a sigma1 (or a sigma2) one.
Surprisingly, BD1047 attenuated the anti-immobility effect of combined treatment with siramesine and MEM (while progesterone did not modify it in a statistically significant manner). These findings are all the more unexpected as neither progesterone nor BD1047 modifies the antidepressant-like effect of joint treatment with SIR and AMA. Elucidation of the above discrepancy requires further studies.
Like in the case of siramesine and AMA, sulpiride and prazosin partly antagonized the antidepressant-like effect of combined treatment with siramesine and MEM, which pointed to a complex mechanism of this interaction, including noradrenergic and dopaminergic mechanisms.
Some results suggested that SA4503 - like DTG or (+)pentazocine -facilitated dopaminergic transmission in the frontal cortex, a brain area believed to play an important role in the pathophysiology of depression (e.g. 33, 34, 35). Biochemical and electrophysiological studies demonstrated that the alterations produced by SA4503 resembled those reported for clinically active ADs (e.g. paroxetine, fluoxetine, citalopram, desipramine) (35). Additionally, our earlier results showed that repeated (but not acute) treatment with SA4503 up-regulated D2-mediated dopaminergic and alpha1-adrenergic transmission, both those effects being important for clinical antidepressant activity (36, 37). According to Rogoz et al. (38), administration of AMA jointly with imipramine induced up-regulation of dopamine D2 and D3 receptors in rat brain: that effect may explain the synergistic action observed in behavioral studies. It should be emphasized that combinations of those drugs, administered according to the same experimental schedule, did not increase locomotor activity.
As mentioned in the Introduction, our earlier studies indicated that ADs potentiated the antidepressant-like effect of uncompetitive NMDA receptor antagonists (13). That interaction was particularly interesting in the case of fluoxetine, which - like siramesine - did not show any antidepressant-like activity in Porsolt's test when it has been given alone (8, 16). Moreover, DTG exerted a synergistic effect with MEM, which was counteracted by progesterone and BD 1047, a sigma antagonist with preferential affinity for sigma1 sites (39).
It is noteworthy that AMA (at therapeutic concentrations), as well as MEM, another uncompetitive NMDA receptor antagonist (at higher doses), bind to sigma sites (40).
In our study, progesterone per se did not modify the immobility time of rats, its anti-immobility effect being also observed, though (e.g. 41). The above discrepancy can be explained by numerous essential procedural differences (e.g. sex, doses and the schedule of administration, modifications in Porsolt's test). The lack of progesterone effect in the forced swim test was reported by other authors (30, 31)
It is well established that all the currently used ADs show therapeutic efficacy in a maximum of 60-70% of depressive patients. The problem of AD-resistant depression has been the subject of an extensive study, and alternative treatments have been proposed (e.g. co-administration of ADs and uncompetitive NMDA receptor antagonists or metyrapone, a glucocorticoid synthesis inhibitor) (16, 42). Preliminary clinical trials concerned with combined administration of imipramine and AMA to patients with unipolar drug-resistant depression are promising (43, 44).
The above-described and some earlier results indicate that the activation of sigma (particularly sigma1) receptor may be one of possible mechanisms by which drugs induce antidepressant-like activity in Porsolt's test, and that this effect may be enhanced by NMDA receptor antagonists. These findings suggest that combined treatment with some sigma ligands and uncompetitive NMDA receptor antagonists may be an alternative to the treatment of antidepressant-resistant depressive patients in the future.
Acknowledgments: The authors wish to express their thanks to Santen Pharmaceutical Co. Ltd. (Japan) for their kind donation of SA4503, and to Lundbeck (Denmark) for siramesine. We are also indebted to Ms. E. Smolak, M.A., for the linguistic supervision of our paper.
- Martin WR, Eades CG, Thompson JA, Huppler RE, Gilbert PE. The effects of morphine- and nalorphine-like drugs in the nondependent and morphine-dependent chronic spinal dog. J Pharmacol Exp Ther 1976; 197: 517-532.
- Seth P, Fei YJ. Li HW, Huang W, Leibach FH, Ganapathy V. Cloning and functional characterization of a sigma receptor from rat brain. J Neurochem 1998; 70: 22-31.
- Quirion R, Bowen W, Itzhak Y, Junien JM, Mussacchio JM, Rothman TP, Su TP, Tam S.W, Taylor DP. A proposal on the classification of sigma binding sites. Trends Pharmacol Sci 1992; 13: 85-86.
- Itzhak Y, Kassim CO. Clorgyline displays high affinity for sigma binding sites in C57BL/6 mouse brain. Eur J Pharmacol 1990; 176: 107-108.
- Largent BL, Wikstrom H, Gundlach AL., Snyder SH.: Structural determinants of s-receptor affinity. Mol Pharmacol, 1987; 32: 772-784.
- Sánchez C, Meier E. Behavioural profile of SSRIs in animal models of depression, anxiety and aggression. Are they all alike? Psychopharmacology 1997; 129: 197-205.
- Matsuno K, Kobayashi T, Tanaka MK, Mita S. s1 Receptor subtype is involved in the relief of behavioral despair in the mouse forced swimming test. Eur J Pharmacol 1996; 312: 267-271.
- Skuza G., Rogó¿ Z.: Effect of combined treatment with selective sigma ligands and amantadine in the forced swimming test in rats. Pol J Pharmacol 2002; 54: 699-702.
- Ukai M, Maeda H, Nanya Y, Kameyama T, Matsuno K. Beneficial effects of acute and repeated administrations of s receptor agonists on behavioral despair in mice exposed to tail suspension. Pharmacol Biochem Behav 1998; 61: 247-253.
- Debonnel G, de Montigny C. Modulation of NMDA and dopaminergic neurotransmissions by sigma ligands: possible implications for the treatment of psychiatric disorders. Life Sci 1996; 58: 721-734.
- Skolnick P. Antidepressants for the new millennium. Eur J Pharmacol 1999; 375: 31-40.
- Willner P. The mesolimbic dopamine system as a target for rapid antidepressant action. Int Clin Psychopharmacol 1997; Suppl 3, S7-S14.
- Maj J, Rogó¿ Z, Skuza G, Sowiñska H. Effects of MK-801 and antidepressant drugs in the forced swimming test in rats. Eur Neuropsychopharmacol 1992; 2: 37-41.
- Papp M, Moryl E. Antidepressant activity of non-competitive and competitive NMDA receptor antagonists in a chronic mild stress model of depression. Eur J Pharmacol 1994; 263: 1-7.
- Trullas R, Skolnick P. Functional antagonists at the NMDA receptor complex exhibit antidepressant actions. Eur J Pharmacol 1990; 185: 1-10.
- Rogozs Z, Skuza G, Maj J, Danysz W. Synergistic effects of uncompetitive NMDA receptor antagonists and antidepressant drugs in the forced swimming test in rats. Neuropharmacology, 2002; 42: 1024-1030.
- Matsumoto RR, Bowen WD, Tom MA, Vo VN, Truong DD, De Costa BR. Characterization of two novel sigma receptor ligands: antidystonic effects in rats suggest sigma receptor antagonism. Eur J Pharmacol 1995; 280: 301-10.
- Maurice T, Roman FJ, Privat A. Modulation by neurosteroids of the in vivo (+)-[3H]SKF-10,047 binding to sigma1 receptors in the mouse forebrain. J Neurosci Res 1996; 46: 734- 743.
- Porsolt RD, Anton G, Blavet N, Jalfre M. Behavioural despair in rats: a new model sensitive to antidepressant treatments. Eur J Pharmacol 1978; 47: 379-391.
- Millan MJ. The role of monoamines in the actions of established and "novel" antidepressant agents: a critical review. European Journal of Pharmacology 2004, 500: 371-384.
- Hayashi T, Su TP. Sigma-1 receptor ligands: potential in the treatment of neuropsychiatric disorders. CNS Drugs 2004; 18: 269-284. Review.
- Hayashi T, Maurice T, Su TP. Ca(2+) signaling via sigma(1)-receptors: novel regulatory mechanism affecting intracellular Ca(2+) concentration. J Pharmacol Exp Ther 2000; 293: 788-798.
- Aydar E, Palmer CP, Klyachko VA, Jackson MB. The sigma receptor as a ligand-regulated auxiliary potassium channel subunit. Neuron 2002; 34: 399-410.
- Matsuno K, Nakazawa N, Okamoto K, Kawashima Y, Mita S. Binding properties of SA4503, a novel and selective s1 receptor agonist. Eur J Pharmacol 1996; 306: 271-279.
- Skuza G, Rogoz Z. Effect of 1,3-di-o-tolylguanidine (DTG), rimcazole and EMD 57445, the s receptor ligands, in the forced swimming test. Pol J Pharmacol 1997; 49: 329-335.
- Akunne HC, Zoski KT, Whetzel SZ, Cordon JJ, Brandon RM, Roman F, Pugsley TA. Neuropharmacological profile of a selective sigma ligand, igmesine: a potential antidepressant. Neuropharmacology 2001; 41: 138-149.
- Tottori K, Miwa T, Uwahodo Y, Yamada S, Nakai M, Oshiro Y, Kikuchi T, Altar CA. Antidepressant-like responses to the combined sigma and 5-HT1A receptor agonist OPC-14523. Neuropharmacology 2001; 41: 976-988.
- Sánchez C, Arnt J, Costall B, Kelly ME, Meier E, Naylor RJ, Perregaard J. The selective s2 ligand Lu 28-179 has potent anxiolytic-like effects in rodents. J Pharmacol Ex Ther 1997; 283: 1323-1332.
- Sánchez C, Papp M. The selective s2 ligand Lu 28-179 has an antidepressant-like profile in the rat chronic mild stress model of depression. Behav Pharmacol 2000; 11: 117-124.
- Maurice T, Urani A, Phan VL, Romieu P. The interaction between neuroactive steroids and the sigma1 receptor function: behavioral consequences and therapeutic opportunities. Brain Res Rev 2001; 37: 116-132.
- Maurice T. Neurosteroids and sigma1 receptors, biochemical and behavioral relevance. Pharmacopsychiatry 2004; 37 Suppl 3: S171-82.
- Couture S, Debonnel G. Modulation of the neuronal response to N-methyl-D-aspartate by selective sigma2 ligands. Synapse 1998; 29: 62-71.
- Gronier B, Debonnel G. Involvement of s receptors in the modulation of the glutamatergic/NMDA neurotransmission in the dopaminergic systems. Eur J Pharmacol 1999; 368: 183-196.
- Gudelsky GA. Effects of s receptor ligands on extracellular concentration of dopamine in the striatum and prefrontal cortex of the rat. Eur J Pharmacol 1995; 286: 223-228.
- Minabe Y, Matsuno K, Ashby CR Jr. Acute and chronic administration of the selective sigma1 receptor agonist SA4503 significantly alters the activity of midbrain dopamine neurons in rats: an in vivo electrophysiological studies. Synapse 1999; 33: 129-140.
- Skuza G, Kolasiewicz W. Repeated treatment with SA4503, a selective sigma1 receptor agonist, up-regulates alpha1-adrenergic system. A behavioral study. Pol J Pharmacol 2001; 53: 547-550.
- Skuza G, Rogoz Z. A potential antidepressant activity of SA4503, a selective s1 receptor agonist. Behav Pharmacol 2002; 13: 537-543.
- Rogó¿ Z, Dlaboga D, Dziedzicka-Wasylewska M. Effect of combined treatment with imipramine and amantadine on the central dopamine D2 and D3 receptors in rats. J Physiol Pharmacol 2003; 54: 257-270.
- Skuza G, Rogoz Z. sigma1 receptor antagonists attenuate antidepressant-like effect induced by co-administration of 1,3 di-o-tolylguanidine (DTG) and memantine in the forced swimming test in rats. Pol J Pharmacol 2003; 55: 1149-1152.
- Kornhuber J, Schoppmeyer K, Riederer P. Affinity of 1-aminoadamantanes for the s binding site in post-mortem human frontal cortex. Neurosci Lett 1993; 163: 129-131.
- Martinez-Mota L, Contreras CM, Saavedra M. Progesterone reduces immobility in rats forced to swim. Arch Med Res 1999; 30:286-289.
- Rogoz Z, Budziszewska B, Kubera M, Basta-Kaim A, Jaworska-Feil L, Skuza G, Lasoñ W. Effect of combined treatment with imipramine and metyrapone on the immobility time, the activity of hypothalamo-pituitary-adrenocortical axis and immunological parameters in the forced swimming test in the rat. J Physiol Pharmacol 2005; 56: 49-61.
- Rogoz Z, Dziedzicka-Wasylewska M, Daniel WA, Wójcikowski J, Dudek D, Wróbel A, Ziêba A. Effects of joint administration of imipramine and amantadine in patients with drug-resistant unipolar depression. Pol J Pharmacol 2004;56: 735-742.
- Stryjer R, Strous RD, Shaked G, Bar F, Feldman B, Kotler M, Polak L, Rosenzcwaig S, Weizman A. Amantadine as augmentation therapy in the management of treatment-resistant depression. Int Clin Psychopharmacol 2003; 18: 93-96.