The deterioration of homeostatic mechanisms that reduces the capability to adapt to changes of internal and external environment seems to represent a hallmark of normal ageing. However, the dissection of physiological "healthy" ageing form changes that accompany pathological ageing processes represents a major difficulty of modern gerontology (1). This dualistic aspect of ageing research becomes obvious when looking at the metabolic and hormonal homeostasis of the elderly. The age-related disregulation of carbohydrate and lipid metabolism is commonly evident as the "epidemy of obesity" and type-2 diabetes. Paradoxically, many elderly suffer from insufficient drive for food intake that results in a decline of lean body mass and may be seen as the "anorexia of ageing" (2). Both sides of these ageing-related disturbances need to be well understood in terms of their pathogenesis to enable better care of the elderly. This review will focus on central regulatory mechanisms of the age-related changes of food intake. It is well known that appetite drive is controlled by both nervous and humoral signals, and that hypothalamus regulates metabolic and energy homeostasis (3). The evidence for the age-related modifications of food intake was obtained mainly from experimental studies performed on laboratory rodents, however, available data on non-human primates and humans will also be presented. While presenting experimenting data attention will be paid to the existence of interstrain variations of metabolic and hormonal indices. Because alterations in the nutritional state, whether short-term or chronic, profoundly affect various aspects of metabolic and hormonal homeostasis (4) the effects of fasting and refeeding on young and aged animals will also be compared.
Main components of central regulatory mechanisms of food intake
It was proposed half a century ago that the regulation of food intake is based
on the activities of two functional hypothalamic centres. On the basis of electrophysiological
observations and lesions of various hypothalamic structures the hunger center,
responsible for the activation of food intake and appropriate behaviour, was
localized to the lateral hypothalamic area (LHA). Because lesions of the VMN
(
ventromedial hypothalamus nucleus) were associated with hyperphagia
and abnormal weight gain, this nucleus was called the "satiety center". VMN
was shown to be activated by the nervous, hormonal and metabolic inputs from
the gastrointestinal tract after commencing food consumption. This elegant hypothesis
was undermined during the last 20 years by the discovery of numerous neuropeptides
and their receptors which were involved in the control of food intake. The distribution
of these peptides throughout CNS was determined, and the electrophysiological
responses of respective brain areas (including NTS,
nucleus tracti solitari,
in the brain stem) to factors known to influence food intake were characterized.
It became obvious, that beside VMN and LHA also other hypothalamic nuclei were
involved in the control of appetite. Moreover, it was proven that hypothalamic
neurons express receptors not only for neuropeptides and neurotransmitters but
also for many hormones and metabolites factors released from peripheral tissues.
Discovering of leptin synthesis in the adipose tissue (5) was a milestone in
the elucidation of pathways used by peripheral hormones and metabolic signals
that affect short- and long-term control of fuel homeostasis. Currently, more
than 50 various factors are known to act at the CNS level to stimulate or inhibit
food intake (6 - 9). Data presented in
Table 1 show that there is a redundancy
of signals that inhibit, and a relative scarcity of signals that stimulate food
intake. In fact, only one peripheral hormone, ghrelin (10), stimulates ingestive
behaviour.
Table 2 shows the distribution of receptors of main factors
involved in the regulation of food intake in major hypothalamic nuclei.
| Table 1.
Neuropeptides and hormones that control food intake |
 |
| Summary of
data reviewed by Cupples (7), Kalra et al. (8), Leibowitz and Wortley
(9). |
| Table 2.
Changes of body weight and food consumption during life span of some rat
strains |
 |
| agrams/100
g body weight; bdecrease of water intake
by 20%. Rat strains: F344, Fischer344; BN, Brown Norway. |
During the last 10 years it was demonstrated that ARC,
hypothalamic arcuate
nucleus (formerly described as
infundibular nucleus) plays a key
role in the control of food intake. ARC embodies neurons that synthesize NPY
(neuropeptide Y) and AgRP (
Agouti-related peptide) which increase food
intake, and another type of cells, described as POMC/
alphaMSH
neurons, that secrete a key anorexigenic peptide, melanocortin (
alphaMSH,
derived from its precursor propetide, POMC). These neurons not only project
to other hypothalamic and CNS nuclei involved in the control of food intake
but also reciprocally inhibit their own activities depending on the kind of
peripheral stimuli that interact with cell receptors (
Fig. 1). Leptin,
insulin, glucose, and cholecystokinin (CCK) belong to the major signals released
in the body in response to food ingestion. These agents are transported into
hypothalamus through the blood-brain-barrier which is less tight in ARC than
in other brain areas (11). Leptin and insulin are major peripheral signals that
simultaneously inhibit NPY/AgRP neurons and stimulate POMC/
alphaMSH
neurons in the ARC (
Fig. 1). In young animals fasting potently up-regulates
hypothalamic mRNA levels of two strong orexigens, AgRP and NPY (5-10 times),
and moderately, by 20-50%, decreases POMC mRNA level (12). These effects are
probably mediated by a substantial decline of blood leptin and insulin levels
that accompanies food deprivation (4, 13).
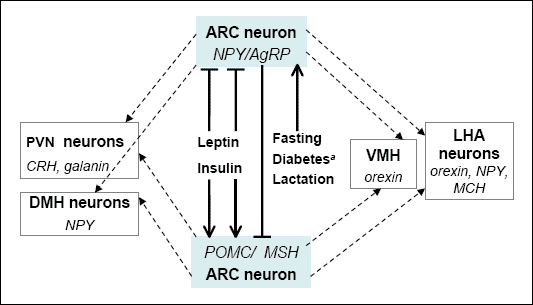 |
| Fig. 1.
Hormonal regulation of hypothalamic appetite centers. |
The activities of neural circuits that control energy homeostasis rely on the delicate balance of factors which finish or stimulate food intake (14, 15). Imbalance of these factors seems to underlie such pathologies as obesity or type 2 diabetes, and may also induce some of the age-related changes of energy homeostasis (16). Because age-related alterations of homeostasis are often revealed upon the stimulation of organ functions (17 - 19) the data on ageing-associated changes of the regulation of fuel metabolism will be reported in the context of fasting and subsequent refeeding, ie. conditions that represent a naturally occurring challenge.
Food consumption in ageing humans and laboratory animals
There is an overwhelming agreement that during normal human ageing there is gradual decline in the amount of food consumed (1, 20). The age-related decrease of food intake is regarded as a normal reaction to the diminished energy expenditure that characterizes healthy elderly. Another feature of normal ageing is the gradual decrease of muscle mass and an increase of fat mass (1). Thus diminished food intake by the elderly may represent physiological adaptation to the decrease in physical activity. However, ageing is associated with important alterations of food perception of that may jeopardize the nutritional status of the elderly. It was shown by Morley and co-workers (2, 20) that many elderly do not consume enough food that leads to the decline of body weight, a dangerous health risk factor. Although the age-related decline of appetite has been known for many years there is little understanding of its physiological basis, especially, in respect to the central regulation of the ingestive behavior.
Because experimental verification of the causes of disturbed food intake can
not be performed in humans answers to this vital problem were sought in animal
experiments in which aged rat represented most commonly used species. However,
in rodents, similarly to humans, there are serious genetic interindividual and
interstrain differences that seem to have a big influence on the control of
energy homeostasis and body mass during ageing. Generally, one can distinguish
two types rat strains with respect to changes of body weight during the life
span. In the first one represented by Wistar and Brown-Norway rats, animals
only modestly increase their weight after reaching maturity, whereas in the
second type rats increase their mass more or less constantly during their life
span (e.g. Sprague-Dawley or Fischer 344). Some rat or mouse strains that were
extensively used in the studies of energy homeostasis cannot be seen as valid
models for ageing research since they show mutated phenotype (eg. Zucker rat,
db/db or
ob/ob mice) that cannot be related to normal ageing process.
Few examples of the differences in food consumption during ageing of various
rat strains are shown in
Table 3.
| Table 3.
Receptors of main neuropeptides and hormones present in rat hypothalamus |
 |
| amost
of POMC neurons co-express also CART immunoreactivity (Cone 2005); bmost
of the NPY neurons co-express also AgRP; cperipheral
ghrelin administration activates NPY/AgRP neurons only in ARC (10). Summary
of data reviewed in (6-12,14-16). |
Age-related differences in the expression and action of neuropeptides and hormones that stimulate or inhibit food intake
NPY
Neuropeptide Y is the best characterized, and probably the most important peptide
that stimulates food intake (3, 8, 9). Therefore, the effect of ageing on the
gene expression of NPY in fed and fasted rats was evaluated by several authors.
Sohn
et al. found that NPY mRNA level was 6 times lower in the hypothalamus
of normal old Brown Norway rats as compared to young ones (21). Similarly, Kaneda
et al. reported that control middle-aged Wistar rats showed 3 times lower
hypothalamic NPY mRNA level than young animals (22).
Fasting prominently increases hypothalamic NPY expression in young animals (12, 23). However, in old rats this adaptory response is strongly suppressed, although genetic variations between rat strains, different age of animals, and duration of fasting period may be a likely cause of differences between studies (23, 24).
Intraventricular (i.c.v.) administration of neuropeptides and hormones is the established method to evaluate their effects on food intake despite the use of high doses of these agents (12). After i.c.v. injection of various doses of NPY virtually no feeding response was observed in senescent rats, in contrast to the dose-dependent high increase of food intake in young animals (24, 25).
The process of ageing is often accompanied by the alterations in the cell and
tissue reactivity to the exogenous agents like drugs or hormones. This may be
caused by the changes in the amount or structure of receptors and signaling
pathways. By using quantitative PCR Coppola
et al. (26) found significant
decline of the mRNA of the Y1 and Y2 NPY receptors in the paraventricular nucleus
of old as compared to young rats. If this finding was confirmed in other hypothalamic
areas and in non-human primates, it would represent a possible mechanism for
the lack of NPY feeding response in old mammals. Paradoxically, the number of
neurons that showed Y1 receptors by immunohistochemistry was twice higher in
old rats as compared to young ones, moreover, the total number of all PVN neurons
did not change with age (26). This age-related increase of NPY type 1 receptors
may compensate for the decreased action of this strong orexigenic neuropeptide.
AgRP
Although many hypothalamic nuclei contain NPY/AgRP neurons (
Table 2)
and both peptides are strong orexigens, their actions are differently affected
by ageing. Similarly to NPY the hypothalamic expression of AgRP is also suppressed
in old rats. However, increase of food intake after IVC administration of AgRP
was well preserved in senescent rats (27). This finding indicates that declined
reactivity to endogenous (like fasting) or exogenous signals (like i.c.v.-administered
agents) did not occur non-specifically but may significantly differ between
various orexigenic signals.
Orexins
Both orexin A (OX-A) and orexin B (OX-B) strongly induce food intake and state of vigilance. It was found that the number of OX-neurons was on average by 28% lower in the lateral hypothalamic area in old versus young rats (28). Whether this decline has functional consequences is controversial. However, i.c.v. administration of orexin A did not stimulate food intake by old rats in contrast to young animals (29). Lack of OX-A action might be caused by the significantly lower amount of type 1 orexin receptors (OX-IR ) in the hypothalamus of old rats (29). However, it was found in the same study that the number of type 2 receptors (OX-2R) was similar in both young and old animals (29).
MSH
In a sharp contrast to NPY, the hypothalamic expression of POMC, the precursor
of anorexigenic
alphaMSH was similar in young
and old rats (30). However, fasting-induced decrease of POMC mRNA was more prominent
in PVN of young as compared to old rats (27).
CART
Although CART, an anorexigenic peptide, is mostly co-localized with POMC in the same neurons, effect of ageing on its expression is quite different. It was demonstrated that the amount of CART mRNA was higher in middle-aged and old rats as compared to young ones in the arcuate nucleus, however, it was lower in the paraventricular nucleus (21, 23).
Leptin
There are numerous data that relate serum leptin concentration to numerous variables.
In ageing either increase or no change in serum leptin levels were reported,
and the variations seem to be related to genetic differences between rat strains
(discussed by Kmiec
et al. in (13)). However, the effect of ageing on
leptin action in the central nervous system was investigated only by few authors
(30). Decreased leptin uptake in hypothalamic nuclei was demonstrated in old
Wistar rats by the use of immunohistochemical methods (31). This decline was
at least partially caused by the diminished expression of the long form of leptin
receptors in the hypothalamus as demonstrated by Western blots and RT-PCR. Interestingly,
despite higher serum leptin level old rats consumed less food than young animals
(31) suggesting age-related leptin resistance.
In conclusion, the presented evidence suggests that, at least in rodents, ageing is associated with the general down-regulation of the expression of hypothalamic peptides that stimulate food intake, and lack of alterations in the hypothalamic expression of anorexigenic peptides. Therefore, in the aged rat the delicate balance that controls food intake between hypothalamic orexigens and anorexigens seems to be shifted to the latter. This situation may be responsible for the decreased appetite drive in senescent animals and loss of weight at their end-of-life period. If the anorexia of ageing observed in humans is based on similar changes at hypothalamus, the therapeutic intervention at this regulatory level would be possible in the future.
Acknowledgements:
This work was supported by the State Committee for Scientific Research Grant
ST-12.
REFERENCES
- Hauser G, Neumann M. Aging with quality of life - a challenge for society. J Physiol Pharmacol 2005; 56 Suppl 2: 35-48.
- Morley JE. Decreased food intake with aging. J Gerontol A 2001; 56A: 81-88.
- Konturek SJ, Konturek JW, Czesnikiewicz-Guzik M, Brzozowski T, Sito E, Konturek PC. Neuro-hormonal control of food intake; basic mechanisms and clinical implications. J Physiol Pharmacol 2005; 56:Suppl 6: 5-25.
- Heber D. Endocrine responses to fasting, malnutrition and illness. In Endocrinology LJ DeGroot (ed) 1977 Philadelphia, WB Saunders: 3: pp. 2663-2685.
- Zhang Y, Proenca R, Maffei M. et al. Positional cloning of the mouse obese gene and its human homologue. Nature 1994; 372: 425-432.
- Schwartz MW, Woods SC, Porte D et al. Central nervous system control of food intake. Nature 2000; 404: 661-671.
- Cupples WA. Physiological regulation of food intake. Am J Physiol Regul Integr Comp Physiol 2005; 288: 1439-1443.
- Kalra SP, Dube MG, Pu S et al. Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr Rev 1999; 20: 68-100.
- Leibowitz SF and Wortley KE. Hypothalamic control of energy balance: different peptides, different functions. Peptides 2004; 25: 473-504.
- Wang L, Saint-Pierre DH, Tache Y. Peripheral ghrelin selectively increases Fos expression in neuropeptide Y-synthesizing neurons in mouse hypothalamic arcuate nucleus. Neurosci Let 2002; 325: 47-51.
- Kastin AJ, Pan W. Dynamic regulation of leptin entry into brain by the blood-brain- barrier. Regulat Pept 2000; 92: 37-43.
- Hakansson ML, Hulting AL, Meister B. Expression of leptin receptor mRNA in the hypothalamic arcuate nucleus - relationship with NPY neurons. Neuroreport 1996; 7: 3087-3092.
- Kmiec Z, Pokrywka L, Kotlarz G, et al. The effects of fasting and refeeding on serum leptin, adiponectin and free fatty acids concentrations in young and old male rats. Gerontology 2005; 51: 357-362.
- Bouret SG and Simerly RB. Leptin and development of hypothalamic feeding circuits. Endocrinology 2004; 145: 2621-2626.
- Cone RG. Anatomy and regulation of central melanocortin system. Nature Neurosci 2005; 8: 571-578.
- Kalra SP, Bagnasco M, Otukonyoung EE, et al. Rhythmic, reciprocal ghrelin and leptin signaling: new insight in the development of obesity. Regul Pept 2003; 111: 1-11.
- Mysliwski A, Kmiec Z. Effect of aging on glycogen synthesis in liver of starved-refed rats. Arch Gerontol Geriatr 1992; 14: 85-92.
- Kmiec Z, Kotlarz G, Mysliwski A et al. Thyroid hormones homeostasis in rats refed after short-term and prolonged starvation. J Endocrinol Invest 1996; 19: 304-311.
- Kmiec Z, Mysliwski A, Wyrzykowska M et al. The effects of fasting and refeeding on serum parathormone and calcitonin concentrations in young and old male rats. Horm Metab Res 2001; 33: 276-280.
- Morley JE, Silver AJ. Anorexia of the elderly. Neurobiol Aging 1988; 9: 9-16.
- Sohn EH, Wolden-Hanson T, Matsumoto AM. Testosterone (T)-induced changes in arcuate nucleus cocaine-amphetamine-regulated transcript and NPY mRNA are attenuated in old compared to young male Brown Norway rats: contribution of T to age-related changes in cocaine-amphetamine-regulated transcript and NPY gene expression. Endocrinology 2002; 143: 954-963.
- Kaneda T, Makino S, Xishiyama M et al. Differential neuropeptide responses to starvation with ageing. J Neuroendocrinol 2001; 13: 1066-1075.
- Gruenewald DA, Marck BT, Matsumoto AM. Fasting-induced increases in food intake and neuropeptide Y gene expression are attenuated in aging male Brown Norway rats. Endocrinology 1996; 137: 4460-4467.
- Blanton CA, Horwitz BA, Blevins JE, et al. Reduced feeding response to neuropeptide Y in senescent Fischer 344 rats. Am J Physiol Regul Integr Comp Physiol 2001; 280: 1052-1060.
- Blanton CA, Horwitz BA, Murtagh-Mark C, et al. Meal patterns associated with the age-related decline in food intake in the Fischer 344 rat. Am J Physiol 1998; 275: 1494-1502.
- Coppola JD, Horwitz BA, Hamilton J, et al. Expression of NPY Yl and Y5 receptors in the hypothalamic paraventricular nucleus of aged Fischer 344 rats. Am J Physiol Regul Integr Comp Physiol 2004; 287: 69-75.
- Wolden-Hanson T, Marck BT, Matsumoto AM. Blunted hypothalamic neuropeptide gene expression in response to fasting, but preservation of feeding responses to AgRP in aging male Brown Norway rats. Am J Physiol Regul Integr Comp Physiol 2004; 287: 138-146.
- Porkka-Heiskannen T, Alanko L, Kalinchuk A, et al. The effect of age on prepro-orexin gene expression and contents of orexin A and B in the rat brain. Neurobiol Aging 2004; 25: 231-238.
- Takano S, Kanai S, Hosoya H, et al. Orexin-A does not stimulate food intake in old rats. Am J Physiol Gastrointest Liver Physiol 2004; 287: 1182-1187.
- Pu S, Dube MG, Kalra PS, et al. Regulation of leptin secretion: effects of aging on daily patterns of serum leptin and food consumption. Reg Pept 2000; 92: 107-111.
- Fernandez-Galaz C, Fernandez-Agullo T, Campoy F, et al. Decreased leptin uptake in hypothalamic nuclei with ageing in Wistar rats. J Endocrinol 2000; 171: 23-32.