As one of the most recently discovered neuropeptides, GAL shows many interesting physiological and behavioral actions, which are mediated through its interactions with distinct G-protein-coupled membrane receptor subtypes (GALR1, GALR2 and GALR3), subsequently activating several signalling pathways (1, 4, 6 - 9). Exogenously administered GAL produces several major biological actions, including inhibitory effects on memory and learning, inhibition of acetylcholine and glutamate release, stimulation of feeding and pituitary hormone release, inhibition of insulin release and spinal nociceptive reflexes (1, 4, 6, 10 - 14). Expression of GAL is regulated by steroids. GAL is dramatically overexpressed after neuronal injury and in Alzheimers disease.
One of the most interesting effect of GAL on the endocrine pancreas is the inhibition of insulin secretion (1 - 3, 5, 8, 10, 11, 15 - 26). In vitro and in vivo studies have shown that GAL (porcine) inhibits insulin secretion in pigs (19), dogs (20), perfused rat pancreas (16, 17, 21, 22), mouse islets (23), rat insulinoma cells (3, 10, 19) and human islets (5, 15). GAL can regulate insulin secretion by direct inhibition of the L-type Ca2+ channels (18, 24) and the activation of the KATP channels (18, 25), important in the control of the ß-cell membrane potential and the influx of Ca2+. The other possible mechanism is the inhibition of adenylate cyclase, leading to the reduction of intracellular Ca2+ concentration (2, 18). Moreover the mechanism of insulin secretion inhibition, that may be exerted by the inhibition of exocytosis at a very late stage in the stimulus-secretion coupling is also suggested (18, 26).
The aim of our present studies is researching effects of new GAL analogues on insulin secretion. We have chosen the 15-amino-acid-residue N-terminal fragment of GAL because this fragment is conserved across species and is recognized by its own receptors. In our studies we have designed and synthesized several GAL analogues modified in positions: 6, 8, 9, 10 and 11 respectively (Table 1). In next step we characterized the biological properties of GAL analogues, investigating their action on glucose-induced insulin secretion from rat isolated islets of Langerhans. We expected, that such modifications may provide more information about molecular mechanisms of peptide-receptor interaction. Research may also lead to finding high-affinity antagonist of GAL receptors.
| Table 1. The primary structures of the synthesized peptides. |
 |
| GAL galanin; Nle norleucine (L-2-aminocaproic acid); Nva norvaline (L-2-aminovaleric acid); Sar sarcosine (N-methylglycine); Tyr(PO3H2) L-phosphotyrosine; |
Synthesis of peptides
All peptides (Table 1) were synthesized by the solid phase peptide synthesis with the use of a Labortec AG model SP 650 peptide synthesizer and 9-fluorenylmethoxycarbonyl (Fmoc) strategy (27). TentaGel S RAM resin for peptide amides (capacity 0.25 mmol/g) was used as the starting material. All amino acids were coupled as active derivatives in a 3-fold molar excess with the use of the O-(benzotriazole-1-yl)-1,1,3,3-tetramethyluronium tetrafluoroborate with addition of N-hydroxybenzotriazole (1:1) coupling method. Deprotection of the Fmoc group was carried out with 20% piperidine in N,N-dimethylformamide. After synthesis had been completed, the peptides were cleaved from resin with trifluoroacetic acid/phenol/triisopropylsilane/water (88:5:2:5 v/v/v/v) mixture. Crude peptides thus obtained were purified by Reverse Phase High Performance Liquid Chromatography (RP-HPLC) on preparative Vydac C18 column (32×240 mm, 15 - 20 µm particle size) with several linear gradients of acetonitrile (ACN) in 0.1% trifluoroacetic acid (TFA). Eluates were fractioned and analyzed by the analytical RP-HPLC. Purity of the peptides (greater than 98%) was checked by an analytical Beckman System Gold chromatograph with a Beckman C18 column (4.6×250 mm, 5 µm particle size) with several linear gradients of ACN in 0.1% TFA. Identities of peptides were confirmed by amino acid analysis and MALDI-TOF mass spectrometry.
Animals and tissue preparation
All procedures were designed in accordance with the generally accepted ethical standards for animal experimentation and accepted by the Local Ethical Committee of the Medical University of Gdañsk. Male Wistar rats (weighting 180 - 220g) were kept under standard laboratory conditions (a natural light-dark cycle) with unrestricted access to food (Labofeed B, Kcynia, Poland) and tap water. Forty eight animals were used in all experiments. Pancreatic rat islets of Langerhans were isolated by collagenase digestion using an intraductal injection technique (28). After the decapitation, a midline laparotomy was performed and 7 - 10 ml of the collagenase type XI solution (1mg/ml) was injected into the duct system of the pancreas. The inflated pancreas was removed from the animal and placed in a water bath at 37°C. Digestion of the pancreas was completed after 10 min and the tissue was rinsed three times with ice-cold Krebs buffer to remove exocrine cells and collagenase. The composition of Krebs buffer (pH 7.38) was (mM): NaCl 120, KCl 4.8, CaCl2 2.5, MgCl2 1.2, NaHCO3 24, bovine serum albumin (0.1%). The islets of comparable size were hand picked under a stereomicroscope and preincubated in Krebs buffer containing 15 mM of glucose. The buffer was bubbled with carbogen (O2/CO2 95/5). Selected islets were allowed to equilibrate for 60 min and then were rinsed with Krebs buffer. Next, they were divided into groups of 3, and each was transferred to separate test tube containing Krebs buffer and placed in a water bath at 37°C.
Determination of insulin secretion
In vitro insulin secretion from the isolated rat pancreatic islets was studied during static incubation. All peptides were tested at two concentrations: 0.1 µM and 1 µM. Each test tube contained Krebs buffer supplemented with glucose at a concentration of 10 mM (control) or 10 mM of glucose with the addition of one peptide tested. The antagonistic/agonistic activities of GAL analogues were checked by the simultaneous application of GAL and one of the galanin analogues, used at the same concentration. The isolated islets were incubated for 60 min in a water bath at 37°C and in the atmosphere of carbogen. After 1h, samples were collected and frozen at 20°C for subsequent analysis of insulin. The concentration of insulin in the incubation medium was determined radioimmunologically (29) using anti-rat insulin antibody and 125I-labelled rat insulin (DRG, USA).
Statistical analysis of the data
Values of insulin level [ng/60min/islet] induced by peptides in the presence of 10 mM glucose are presented as means±SEM (standard error of mean). Students t-test and one-way ANOVA were used to test the degree of significance. Two tailed P values of less than 0,05 were interpreted as indicating a significant difference.
Porcine GAL, GAL(1-15)NH2 and its twelve new analogues modified in positions 6, 8, 9, 10 and 11 were tested for their effect on glucose-induced insulin secretion from rat pancreatic islets of Langerhans. Our studies have shown that GAL and its analogues modulate glucose-induced insulin release from rat pancreatic islets in a concentration-dependent manner. Porcine GAL and its fragment GAL(1-15)NH2 at all concentration tested inhibit glucose-induced insulin secretion. However the N-terminal fragment of GAL inhibited insulin secretion with lower potency in comparison to native GAL.
 |
| Figure 1. Effects of GAL(1-15) analogues modified in positions 6 and 8 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM glucose. |
We have shown that the substitution of Ser6 in the N-terminal part of GAL(1-15) fragment with its stereoisomer, D-Ser, results in increase of the affinity towards GAL receptors. Analogue [D-Ser6]GAL(1-15)NH2 lowered the glucose-induced insulin level by about 63% at a concentration of 0.1 µM and by about 61% at a concentration of 1 µM. It also significantly increased the inhibitory action of GAL even repeatedly at both concentrations used. The simultaneous substitution of Ser6 with D-Ser and Gly8 with D-Trp in GAL(1-15) structure also caused a decrease in the insulin secretion by about 40% and 47% (at a lower and higher concentration, respectively). The simultaneous application of analogue [D-Ser6,D-Trp8]GAL(1-15)NH2 and GAL (at the same concentrations) caused an increase in the inhibitory action of GAL, similar to that for analogue [D-Ser6]GAL(1-15)NH2. In this case we observed repeated increase in inhibitory activity of GAL. The substitution of Gly8 in GAL(1-15) structure with its N-methylated form, sarcosine, also caused an increase in the affinity towards GAL receptors. The analogue [Sar8]GAL(1-15)NH2 showed the inhibitory activity on glucose-induced insulin secretion as it lowered the insulin level by about 21% and 39% (at a lower and higher concentration, respectively). It also significantly increased the inhibitory action of GAL. Similarly like two mentioned above analogues, it extended repeatedly inhibitory action of GAL at both concentrations used.
 |
| Figure 2. Effect of GAL1-15) analogue modified in position 9 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM glucose. |
We showed that the substitution of Tyr9 with Phe caused a substantial change in the galanin effect on glucose-induced insulin secretion. Analogue [Phe9]GAL(1-15)NH2 showed insulinotropic activity as compared to GAL or GAL(1-15)NH2. It caused an increase in the insulin secretion by about 66% and 48% at a concentration of 1 µM and 0.1 µM, respectively. Our studies have shown that it completely abolished inhibitory effect of GAL (when simultaneously used with GAL at the same concentrations) and still showed the insulinotropic activity. However the replacement of Tyr9 in GAL(1-15) structure with its phosphorylated form, analogue [Tyr(PO3H2)9]GAL(1-15)NH2, affects the glucose-induced insulin secretion only at a higher concentration. It caused an increase in the insulin level by about 12%. This peptide used at a both concentrations tested abolished inhibitory action of GAL, although its effect was stronger at higher concentration. Also the analogue [Trp9]GAL(1-15)NH2, in which Tyr9 was replaced with Trp, another aromatic residue, showed insulinotropic activity as compared to GAL or GAL(1-15)NH2. It caused an increase in the insulin secretion by about 15% and 29% (at a lower and higher concentration, respectively). This peptide, used at a concentration of 0.1 µM, also reduced inhibitory effect of GAL by about 14%, but at a concentration of 1 µM completely abolished the inhibitory action of GAL.
 |
| Figure 3. Effects of GAL(1-15) analogues modified in positions 9 and 10 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM glucose (*P<0.05 vs. GAL). |
The insulinotropic activity also showed the analogue GAL(1-15) in which Leu10 was replaced with its D-stereoisomer. Analogue [D-Leu10]GAL(1-15)NH2 used at a concentration of 1 µM caused an increase in the insulin secretion by about 29% and by about 16% at a concentration of 0.1 µM. Moreover this peptide used at a concentration of 0.1 µM reduced the inhibitory effect of GAL and at higher concentration slightly stimulated secretion of insulin by about 14%. The substitution of Leu10 with its homologue, Val, gave in consequence the peptide which stimulated the insulin secretion in the tests on glucose-induced insulin secretion from isolated rat pancreatic islets of Langerhans. Analogue [Val10]GAL(1-15)NH2 increased the insulin level by about 25% and 31% (at a concentration of 0.1 µM and 1 µM, respectively). Simultaneously this peptide abolished the inhibitory action of GAL and slightly increased the insulin level by about 17% and 13% (at a lower and higher concentration, respectively). Whereas the analogue, in which Leu10 was substituted with Nva have partly shown the insulinotropic activities, only at a concentration of 0.1 µM. Analogue [Nva10]GAL(1-15)NH2 completely abolished the inhibitory action of GAL at both concentrations used.
 |
| Figure 4. Effects of GAL(1-15) analogues modified in positions 9 and 10 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM glucose in the presence of appropriate concentration of GAL. |
The lack of inhibition of insulin secretion was observed in the case of analogue in which Leu11 was replaced with Pro. This peptide increased insulin secretion by about 21% (at lower concentration) and 26% (at higher concentration). Moreover the analogue [Pro11]GAL(1-15)NH2 showed strong antagonistic activity. At the both concentrations used, it completely abolished the inhibitory action of GAL. Two another analogues, in which Leu11 was substituted with its structural isomer Ile or Nle, caused a decrease in the insulin secretion by about 19% (at a concentration of 0.1 µM) or 26% (at a concentration of 1 µM) and by about 14% (at a concentration of 0.1 µM) and 20% (at a concentration of 1 µM), respectively. Whereas analogues: [Ile11]GAL(1-15)NH2 and [Nle11]GAL(1-15)NH2 only slightly reduced the GAL activity with a lower potency than [Pro11]GAL(1-15)NH2.
 |
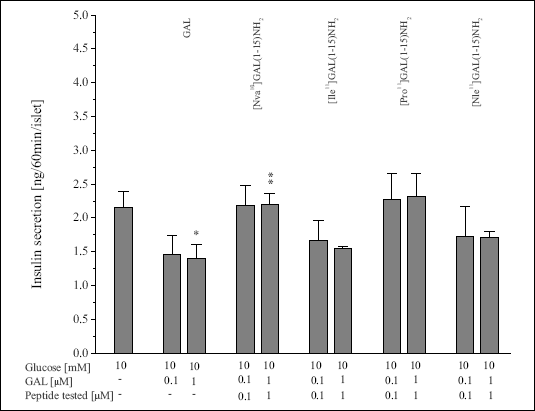
| Figure 5. Effects of GAL(1-15) analogues modified in positions 10 and 11 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM glucose (*P<0.05 vs. 10 mM glucose, **P<0.05 vs. GAL). |
 |
| Figure 6. Effects of GAL(1-15) analogues modified in positions 10 and 11 on insulin secretion from the rat pancreatic islets of Langerhans induced by 10 mM in the presence of appropriate concentration of GAL (*P<0.05 vs. 10 mM glucose, **P<0.05 vs. GAL). |
Galanin is a 29/30 amino-acid-residue neuropeptide which is implicated in the treatment of Alzheimers disease, depression, feeding disorders, prevention of ischaemic damage and the treatment of chronic pain (1, 4, 6, 12 - 14, 30 - 37). The N-terminus of GAL is highly conserved between different species, and the first 15 amino acids were found to be sufficient for agonistic receptor binding. Full-length GAL has a high cross-reactivity towards receptors from other species with similar affinity, although the agonistic effect may not be preserved (4, 6 - 10, 13). It has been shown that the one of major action of GAL is inhibition of insulin secretion (1 - 3, 5, 6, 15 - 26). GAL decrease adenylate cyclase activity and intracellular cAMP production via a pertussis-toxinsensitive pathway, involving an inhibitory GTP-binding protein (2, 18). In addition, GAL was reported to activate ATP-sensitive potassium channels in ß-cells (18, 25).
Nowadays numerous GAL fragments, chimeras and analogues with point mutation were synthesized and their activities on the insulin secretion were studied (3, 10, 11, 17, 21, 22, 30 - 33, 38 - 45). Structure-activity studies have shown that the activity of GAL is connected with the N-terminal 1-15 fragment, needed for full activation of GAL receptors and the inhibitory effect of GAL on insulin secretion. The study with the use of GAL analogues indicate that Gly1 and Trp2 in GAL structure are of great importance in the binding to GAL receptors (7 - 9). It is known that GAL(2 - 29) fragment have substantially lower affinity to GAL receptors than does full-length GAL molecule. Trp2 residue is the most important pharmacophore in GAL structure and it is though to interact with a pair of histidines placed in trans-membrane domain VI of the GALR1 receptor (7). All attempts to modify or replace the indole structure of Trp have resulted in analogues with very low or lack of affinity. The important pharmacophores are also Asn5 , Tyr9, Leu10 and Leu11 (13, 39).
In our studies we have designed twelve new GAL analogues modified in positions: 6, 8, 9, 10, 11 and studied their action on glucose-induced insulin secretion. Our observations confirmed that GAL and its N-terminal fragment GAL(1-15)NH2 are full GAL receptor agonists in the isolated rat pancreatic islets of Langerhans. They lowered glucose-induced insulin secretion, but fragment GAL(1-15)NH2 was about 30% less potent inhibitor than GAL. It suggests that the C-terminal fragment GAL 16-29 is required for full activations of GAL receptors in the rat pancreatic islets.
However studies with the use of the GAL(1-15)NH2 analogues have shown diverse activities of such modified peptides as compared to GAL or its non-modified N-terminal fragment 1-15. Analogues: [D-Ser6]GAL(1-15)NH2, [D-Ser6,D-Trp8]GAL(1-15)NH2 and [Sar8]GAL(1-15)NH2 have been found to be very strong GAL receptor agonists. They lowered the insulin level in the tests on glucose-induced insulin secretion and significantly increased the inhibitory effect of GAL. Two other analogues: [Nle11]GAL(1-15)NH2 and [Ile11]GAL(1-15)NH2 also showed the agonistic activities, but they were less active than mentioned above three peptides.
More significant changes in the biological activity of GAL(1-15)NH2 resulted from modifications of the amino acid sequence of GAL, in which Tyr9 was replaced by Tyr(PO3H2) or Trp and Leu10 was substituted with D-Leu, Val or Nva. Such modified GAL analogues used at a concentration of 1 µM, were able to stimulate the insulin secretion from rat pancreatic islets with the following order of insulinotropic potency: [Trp9]GAL(1-15)NH2
We have shown that the strongest antagonistic properties owned two GAL analogues: [Phe9]GAL(1-15)NH2 and [Pro11]GAL(1-15)NH2. They stimulated glucose-induced insulin secretion and antagonized the inhibitory effect of GAL in a concentration-dependent manner. Relatively the strongest antagonistic activities showed the analogue [Phe9]GAL(1-15)NH2 used at a concentration of 1 µM. Our observations showed that the modification of the amino acid sequence of GAL(1-15)NH2, in which Tyr9 was replaced with aromatic ring of Phe is the most advantageous for its antagonistic properties. We suggest that aromatic phenyl, not hydroxyl, group is involved in the interaction with GAL receptors.
In our studies we showed that residues in positions: Ser6, Gly8, Tyr9, Leu10 and Leu11 within GAL structure may play an important role in the interaction of GAL with its receptors presented in rat pancreas. The modifications of GAL structure in positions 6 and/or 8 resulted in increase of the affinity towards GAL receptors and led to finding analogues with stronger agonistic activities than non-modified GAL(1-15) fragment. However the modifications of GAL structure in positions 9, 10 or 11 gave in consequence peptides which showed insulinotropic activities and antagonized the inhibitory action of GAL in a concentration-dependent manner. We expect that these findings provide more information about molecular mechanisms of galanin-receptor interaction and may be a useful tool for developing a new more potent GAL receptor antagonists or agonist in pancreas.
Ackonowledgments: This work was supported by EFS and grant no. BW-8000-5-0400-7 from the University of Gdansk.
- Crawley JN. Biological actions of galanin. Regul Peptides 1995; 59: 1-16.
- Amiranoff B, Lorinet AM, Lagny-Pourmir I, Laburthe M. Mechanism of galanin-inhibited insulin release. Occurrence of a pertussis-toxin-sensitive inhibition of adenylate cyclase. Eur J Biochem 1988; 177: 147-152.
- Amiranoff B, Lorinet AM, Yanaihara N, Laburthe M. Structural requirement for galanin action in the pancreatic beta cell line RINm5F. Eur J Pharmacol 1989; 163: 205-207.
- Bartfai T, Fisone G, Langel U. Galanin and galanin antagonists: molecular and biochemical perspectives. Trends Pharmacol Sci 1992; 13: 312-317.
- Dunning B, ArRajab A, Bottcher G, Sundler F, Ahren B. Galanin (GAL) is present in human pancreatic nerves and inhibits insulin release (IRI) from isolated human islets. Diabetes 1990; 39: 135A.
- Kask K, Langel U, Bartfai T. Galanin - a neuropeptide with inhibitory actions. Cell Mol Neurobiol 1995; 15: 653-673.
- Floren A, Land T, Langel U. Galanin receptor subtypes and ligand binding. Neuropeptides 2000; 34: 331-337.
- Lu X, Lundström L, Langel Ulo, Bartfai T. Galanin receptor ligands. Neuropeptides 2005; 39: 143-146.
- Lundström L, Lu X, Langel Ü, Bartfai T. Important pharmacophores for binding to galanin receptor 2. Neuropeptides 2005; 39: 169-171.
- Gallwitz B, Schmidt WE, Schwarzhoff R, Creuztzfeldt W. Galanin: structural requirements for binding and signal transduction in RINm5F insulinoma cells. Biochem Biophys Res Commun 1990; 172: 268-275.
- Kakuyama H, Mochizuki T, Iguchi K, et al. [Ala6,D-Trp8]-galanin(1-15)ol is a potent galanin antagonist on insulin release. Biomed Res 1997; 18: 49-56.
- Kuteeva E, Wardi T, Hökfelt T, Ögren SO. Galanin enhances and a galanin antagonist attenuates depression-like behaviour in the rat. Eur Neuropsychopharmacol 2007; 17: 64-69.
- Land T, Langel U, Low M, Berthold M, Unden A, Bartfai T. Linear and cyclic N-terminal galanin fragments and analogs as ligands at the hypothalamic galanin receptor. Int J Peptide Prot Res 1991; 38: 267-272.
- Xu ZQD, Zheng K, Hökfelt T. Electrophysiological studies on galanin effects in brain progress during the last six years. Neuropeptides 2005; 39: 269-275.
- Gilbey SG, Stephenson J, OHalloron DJ, Burrin JM, Bloom SR. High-dose porcine galanin infusion and effect on intravenous glucose tolerance in humans. Diabetes 1989; 38: 1114-1116.
- Leonhardt U, Siegel EG, Kohler H, et al. Galanin inhibits glucose-induced insulin release in vitro. Hormone Metab Res 1989; 21: 100-101.
- Ruczynski J, Cybal M, Rekowski P, Wójcikowski C. Effects of porcine galanin, galanin(1-15)NH2 and its new analogues on glucose-induced insulin secretion. Pol J Pharmacol 2002; 54: 133-141.
- Sharp GW. Mechanisms of inhibition of insulin release. Amer J Physiol 1996; 271: C1781-1799.
- Lindskog S, Dunning BE, Martensson H, ArRajab A, Taborsky GJ, Ahrén B. Galanin of the homologous species inhibits insulin secretion in the rat and in the pig. Acta Physiol Scand 1990; 139: 591-596.
- McDonald TJ, Dupre J, Tatemoto K, Greenberg GR, Radziuk J, Mutt V. Galanin inhibits insulin secretion and induces hyperglycemia in dogs. Diabetes 1985; 34: 192-196.
- Rekowski P, Borowiec A, Druzynska J, Kusiak E. New chimeric analogues of galanin: synthesis and effects on the glucose-induced insulin secretion. Lett Pept Sci 1998; 5: 417-420.
- Ruczynski J, Konstanski Z, Cybal M, et al. Synthesis and biological properties of new chimeric galanin analogue GAL(1-13)-[Ala10,11]ET-1(6-21)-NH2. J Physiol Pharmacol 2005; 56: 273-285.
- Lindskog S, Ahrén B. Galanin: effects on basal and stimulated insulin and glucagon secretion in the mouse. Acta Physiol Scand 1987; 129: 305-309.
- Homaidan FR, Sharp GW, Nowak LM. Galanin inhibits a dihydropyridine-sensitive Ca2+ current in the RINm5f cell line. Proc Nat Acad Sci USA 1991; 88: 8744-8748
- Straub SG, James RF, Dunne MJ, Sharp GW. Glucose activates both KATP channel-dependent and KATP channel-independent signaling pathways in human islets. Diabetes 1998; 47: 758-763.
- Sharp GW, Le Marchand-Brustel Y, Yada T, et al. Galanin can inhibit insulin release by a mechanism other than membrane hyperpolarization or inhibition of adenylate cyclase. J Biol Chem 1989; 264: 7302-7309.
- Chan WC, White PD. Fmoc Solid Phase Peptide Synthesis. A Practical Approach. Oxford, Oxford University Press, 2000.
- Lacy PE, Kostianovsky M. Metod for the isolation of intact islets of Langerhans from the rat pancreas. Diabetes 1967; 16: 35-39.
- Morgan CR, Lazarow A. Immunoassay of insulin: two antibody system. Plasma insulin levels of normal, subdiabetic and diabetic rats. Diabetes 1963; 12: 115-120.
- Mochizuki T, Ishikawa J, Takatsuka N, Hoshino M, Yanaihara N. Structure-function relationship of galanin. Proc 28th Symp Pept Chem, Osaka, 1990, Ed. Shimonishi Y, Protein Research Foundation, Osaka, 373-377, 1991.
- Mochizuki T, Sato T, Takatsuka N, et al. Synthesis of galanin antagonist. Proc 31st Symp Pept Chem, Osaka, 1993, Ed. Okada Y, Protein Research Foundation, Osaka, 325-328, 1994.
- Hökfelt T. Galanin and its receptors: Introduction to the Third International Symposium, San Diego, California, USA, 2122 October 2004. Neuropeptides 2005; 39: 125-142.
- Lang R, Gundlach A, Kofler B. The galanin peptide family: Receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol Ther 2007; 115: 177-207.
- Lu X, Sharkey L, Bartfai T. The brain galanin receptors: targets for novel antidepressant drugs. CNS Neurol Disord Drug Targets 2007; 6: 183-192.
- Brewer A, Echevarria DJ, Langel Û, Robinson JK. Assessment of new functional roles for galanin in the CNS. Neuropeptides 2005; 39: 323-326.
- Brumovsky P, Mennicken F, ODonnell D, Hökfelt T. Differential distribution and regulation of galanin receptors-1 and -2 in the rat lumbar spinal cord. Brain Res 2006; 1085: 111-120.
- Zorrilla EP, Brennan M, Sabino V, Lu X, Bartfai T. Galanin type 1 receptor knockout mice show altered responses to high-fat diet and glucose challenge. Physiol Behav 2007; 91: 479-485.
- Lagny-Pourmir I, Lorinet AM, Yanaihara N, Laburthe M. Structural requirements for galanin interaction with receptors from pancreatic beta cells and from brain tissue of the rat. Peptides 1989; 10: 757-761.
- Yanaihara N, Mochizuki T, Takatsuka N, et al. Galanin analogues: agonist and antagonist. Regul Peptides 1993; 46: 93-101.
- Pooga M, Jureus A, Rezael K, et al. Novel galanin receptor ligands. J Peptide Res 1998; 51: 65-74.
- Ruczyñski J, Konstañski Z, Korolkiewicz R, Petrusewicz J, Rekowski P. Galanin and its analogues: a structure-activity relationship studies in rat isolated gastric smooth muscles. Lett Pept Sci 2002; 9: 91-99.
- Lu X, Lundström L, Langel U, Bartfai T. Galanin receptor ligands. Neuropeptides 2005; 39: 143-146.
- Anselmi L, Salvatore S, Lakhter A, Hirano A, Tonini M, Sternini C. Galanin receptors in the rat gastrointestinal tract. Neuropeptides 2005; 39: 349-352.
- Legakis IN, Mantzouridis T, Mountokalakis T. Positive correlation of galanin with glucose in healthy volunteers during an oral glucose tolerance test. Horm Metab Res 2007; 39: 53-55.
- Anselmi L, Lakhter A, Hirano A, Tonini M, Sternini C. Expression of galanin receptor messenger RNAs in different regions of the rat gastrointestinal tract. Peptides 2005; 26: 815-819.